
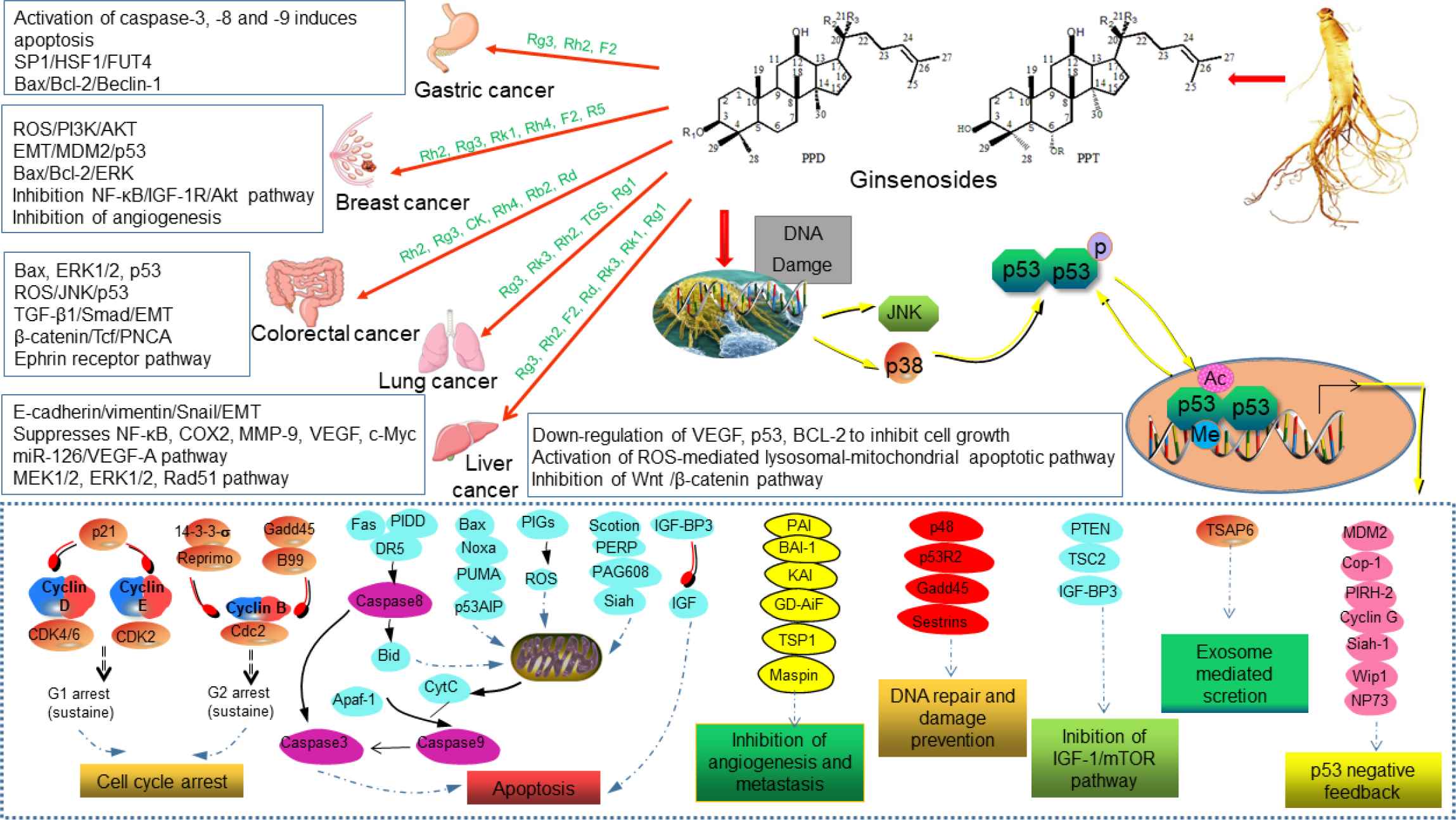
The Anticancer Activity and Mechanisms of Ginsenosides: An Updated Review
- DOI
- 10.2991/efood.k.200512.001How to use a DOI?
- Keywords
- Ginseng; ginsenosides; biological activity; cancer therapy; mechanism
- Abstract
The annual global mortality rate of cancer has increased dramatically. Although various therapies are used for cancer, the results have not been satisfactory. Chemotherapy is currently the most common treatment option. However, serious side effects and drug resistance impede the therapeutic efficacy of chemotherapeutic drugs. Increasing evidence has shown that ginsenosides as a type of phytochemicals play an important role in the prevention and treatment of cancers, such as colon cancer, cervical cancer, ovarian cancer, breast cancer, lung cancer, pancreatic cancer, bladder cancer, esophageal cancer, and bone cancer. Ginsenosides block a variety of enzymes required for tumor growth, which regulate an array of cell progression, such as nitric oxide synthase activity, protein kinase activity, epidermal growth factor receptor intrinsic kinase activity, and nuclear factor-kappaB activity. Ginsenosides also inhibit lipid peroxidation and the production of reactive oxygen species. Thus, ginsenosides can be used as an adjuvant to conventional cancer therapies to improve efficacy and/or reduce side effects through synergistic activity. In this review, we summarized the latest research advances of the anticancer effects of ginsenosides and their potential mechanisms.
- Graphical Abstract
- Copyright
- © 2020 International Association of Dietetic Nutrition and Safety. Publishing services by Atlantis Press International B.V.
- Open Access
- This is an open access article distributed under the CC BY-NC 4.0 license (http://creativecommons.org/licenses/by-nc/4.0/).
1. INTRODUCTION
Over the past few decades, researcher has revealed that the occurrence and progression of most human cancers involve certain molecular biology, biochemical, and cell biology principles. However, the occurrence of human cancer is a complicated process. The genetic variation of certain cells and genes drives normal cells to become highly malignant cancer cells [1]. Pathological analysis of a variety of organ tissues seems to reveal an intermediate step, which participates in the gradual evolution of the normal state to an invasive cancerous state through a series of precancerous conditions [2]. Observations of cancer models in humans and animals suggest that tumor progression involves a range of genetic changes, each of which confers one or more growth advantage. Due to the defect of the regulatory loop that maintains normal cell proliferation and homeostasis, resulting in the normal cells gradually transform into cancer cells in the tumor microenvironment.
The development of human tumors usually involves several hallmark biological capabilities: self-sufficiency in growth signals, insensitivity to growth inhibition (anti-growth) signals, a program to circumvent cell death (apoptosis), infinite replication potential, sustained angiogenesis, tissue invasion and metastasis, energy metabolism reprogramming, and the ability to evade immune destruction [3,4]. Together, these changes determine malignant growth. Tumors are not only island-like masses that proliferate cancer cells, but also are complex tissues composed of many different cell types that participate in each other’s heterotypic interactions. As normal cells gradually evolve into tumor states, they acquire the continuity of these iconic abilities. Cancer is the first or second leading cause of death before age 70 in 91 of 172 countries (WHO) [5]. According to the latest statistics from the Global Cancer Observatory in 2018 (http://gco.iarc.fr/today), the proportion of cancers with higher incidence and mortality estimated as shown in Figure 1A, indicating that the incidence of lung cancer, breast cancer, colorectal cancer, pancreas cancer, liver cancer, stomach cancer, and esophagus cancer is 11.6%, 11.6%, 10.2%, 7.1%, 4.7%, 5.7%, and 3.2%, and the corresponding mortality are 6.6%, 18.4%, 9.2%, 4.5%, 8.2%, 8.2%, and 5.3%; among the continents, Asia is hardest hit affected region for cancer with an estimated 8.7 million cases and 0.055 million deaths, followed by Europe, North America, Latin America & Caribbean, and Africa, and Oceania is the least affected (Figure 1B).
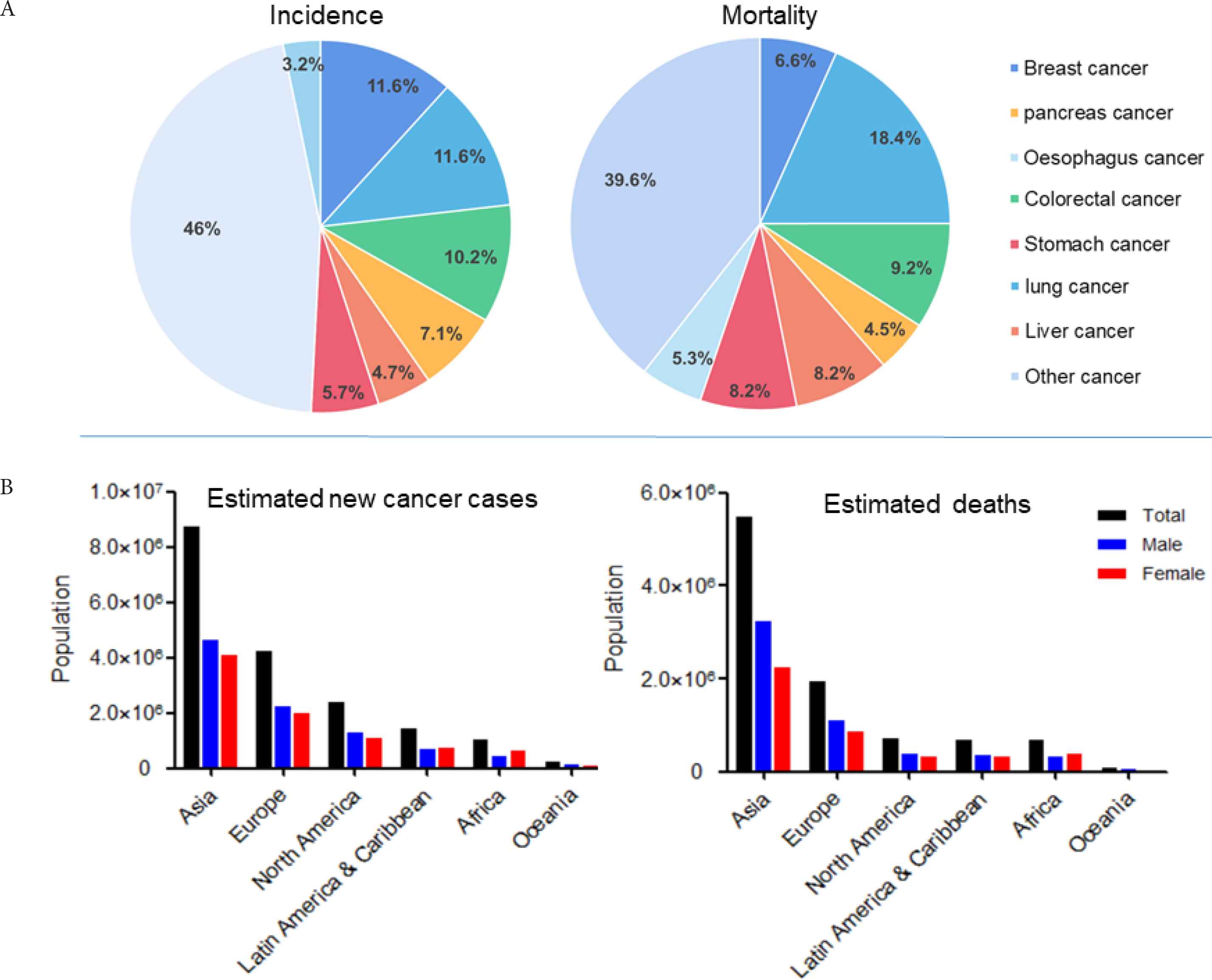
The global distribution of cancer in 2018 (Data from the global cancer observatory). (A) The incidence and mortality of different cancers. (B) The estimated new cancer cases and deaths by sex in every continent of the world.
Cancer treatment is generally divided into drug therapy and non-drug therapy, of which chemotherapy is an important means of drug therapy. Tumor chemotherapy generally uses chemical drugs called cytotoxic drugs, with high side effect rates that cause very serious damage to normal cells [6,7]. Chemotherapy, surgery, and radiotherapy are the three major treatments for cancer and some cancer patients can achieve clinical cures or long-term survival through chemotherapy. Cisplatin exhibits an anti-lung tumor effect by impairing the structure and function of DNA, and pemetrexed regulates intracellular folate-dependent metabolism and inhibits cancer cell replication and proliferation [8]. Erlotinib inhibits tumor growth by targeting Epidermal Growth Factor (EGFR) to inhibit EGFR and its downstream signaling pathways [9]. However, clinical data indicate that these drugs have limited application at tolerated doses due to their toxic side effects and drug resistance [10]. Although chemotherapy has made great progress in the past few decades, targeted drugs have been developed in recent years. However, side effects and drug resistance have caused the most patients to die from cancer invasion and metastasis, with a 5-year survival rate of <15% [7]. Natural products have become a major resource for the development of anticancer agents due to their biochemical activity. Several plant-derived compounds, such as vinblastine, etoposide, paclitaxel, vinorelbine, and irinotecan, have been approved for clinical anticancer drugs. More than 35,000 plants in different countries are used for medicinal purposes around the world [11]. Almost 80% of the world’s population has primary healthcare that relies on traditional medicine, most of which involve the consumption of plant extracts [11]. Natural products can be used as the main source for screening of the targeted anticancer drugs due to the low toxicity and side effects.
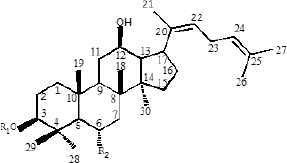
Ginseng (Panax ginseng C.A. Mey) is a valuable oriental Chinese herbal medicine. Because of its effective medicinal activity, it has received increasing attention for the treatment of cardiovascular disease, diabetes, and central nervous system diseases [12–14]. Ginsenoside is an active pharmaceutical ingredient extracted from the traditional Chinese herb ginseng and is widely used for its anticancer [15], apoptosis [16], angiogenesis [17], oxidative stress [18], inflammation [19], and cancer metastasis [20] activities related to cell proliferation. Currently, 40 types of ginsenosides have been identified and most ginsenosides have a dammarane triterpenoid structure [21]. Differences in sugar types, the number of glycosyl groups, and the attachment position provide a diversity of ginsenoside structures. According to the presence of hydroxyl groups at C-6, ginsenosides can be divided into two groups, Protopanaxadiols (PPD) (including Rb1, Rd, Rh2, Rg3, Rg5, and Rk1) and Protopanaxatriols (PPT) (including Rg1, Rh1, RK3, and Rh4) [21]. Studies have shown that the steaming process results in the conversion of large amounts of ginsenosides in ginseng to new degradants, including Rh3, Rk3, Rh4, Rk1, Rk2, and Rg5 (Table 1), which are rare ginsenosides with significant chemo-preventive effects [22].
| Structure skeletons | Ginsenoside | R1 | R2 | R3 |
|---|---|---|---|---|
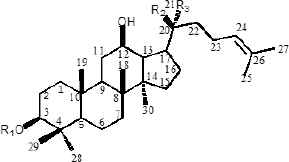 |
20(S)-Rg3 | glc(2-1)glc | OH | CH3 |
| 20(R)-Rg3 | glc(2-1)glc | CH3 | OH | |
| Rb1 | glc(2-1)glc | Oglc(6-1)glc | CH3 | |
| Rb2 | glc(2-1)glc | Oglc(6-1)arap | CH3 | |
| Rh2 | glc | CH3 | OH | |
| Rc | glc(2-1)glc | Oglc(6-1)araf | CH3 | |
| Rd | glc(2-1)glc | Oglc | CH3 | |
| Compound K | H | Oglc | CH3 | |
| PPD | H | OH | CH3 | |
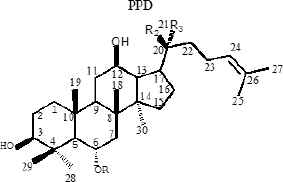 |
Rg1 | glc | Oglc | CH3 |
| Rg2 | glc(2-1)rha | OH | CH3 | |
| 20(S)-Rh1 | glc | OH | CH3 | |
| 20(R)-Rh1 | glc | CH3 | OH | |
| Re | glc(2-1)rha | Oglc | CH3 | |
| Rf | glc(2-1)glc | OH | CH3 | |
| F1 | H | Oglc | CH3 | |
| PPT | H | OH | CH3 | |
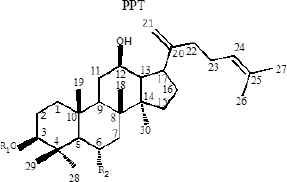 |
Rk3 | H | Oglc | — |
| Rk1 | glc(2-1)glc | H | — | |
| Rk2 | glc | H | — | |
 |
Rh4 | H | Oglc | — |
| Rg5 | glc(2-1)glc | H | — | |
| Rh3 | glc | H | — |
Glc, β-
Structures of protopanaxadiols (PPD), protopanaxatriols (PPT), and rare ginsenosides
To systematically clarify the anticancer activity and mechanisms of ginsenosides, scientists have done many in-depth studies using in vivo and in vitro models over the past few decades. A comprehensive review [23] indicated that ginsenosides are a special molecule in many other naturally occurring cancer therapy compounds. The pleiotropic nature of sapogenin allows it to target the genome (DNA), messengers (RNA), and enzymes (protein) within the cell. Unlike other chemotherapeutic drugs, ginsenosides are pleiotropic and involve in Nuclear Factor-kappaB (NF-κB), Transcription Factor (TF), Activated Protein-1 (AP-1), tumor protein 53 (p53), and cell–cell adhesion molecules, Mitogen-activated Protein Kinase (MAPK), and nuclear β-catenin signaling. In this report, the available literature for the past 10 years was reviewed and discussed to facilitate further research of ginsenosides in cancer.
2. ANTICANCER ACTIVITY AND MECHANISMS
2.1. Breast Cancer
Globally, metastatic breast cancer is the most common cause of female death in recent years and one of the most common malignant tumors [24]. Chemotherapy, hormonal therapy, radiation therapy, and limited or radical surgery are treatments used for breast cancer. The surgical treatment of breast cancer includes Breast-conserving Surgery (BCS) or mastectomy [25]. Local tumors are treated with radiation therapy after BCS, with total long-term patient survival similar to that achieved by mastectomy. Rh2 inhibits the growth of MCF-7 breast cancer cells by inducing the expression of cyclin p21 and decreasing the level of cyclin D3, leading to the up-regulation of Cyclin-dependent Kinase (Cdk) inhibitor p21WAF1/CIP1, which reduces the binding of phosphorylated retinoblastoma protein and transcription factor E2F-1 [26]. In addition, p15 INK4B and p27 KIP1-dependent kinase activities are overexpressed in breast cancer cells and Rh2 regulatory kinase activity induces the cell cycle arrest of breast cancer cell lines in the G1 phase, lowering the viability of MCF-7 and MDA-MB-231 breast cancer cells [27]. Rg3 inhibits NF-κB signaling by regulation of Bax/Bcl-2 expression in triple negative breast cancer (TNBC) and promotes cytotoxicity and the apoptosis of TNBC cells and xenografts in response to treatment with paclitaxel [28]. Apoptosis is thought to be the primary mechanism by which chemotherapeutic agents kill cells. It is a highly conserved form of programmed cell death that regulates tissue homeostasis and/or eliminates damaged and infected cells. There are two major apoptotic pathways, the extrinsic pathway mediated by death receptors and the mitochondria-mediated intrinsic pathway [29]. In the MDA-MB-231 cell line, Rg3 suppresses the phosphorylation of Extracellular Signal-regulated Kinase (ERK) and protein kinase B (Akt), thereby preventing the activation of NF-κB to cause apoptosis [30]. Our group found that Rk1 induces cell cycle arrest and apoptosis in MDA-MB-231 triple negative breast cancer cells by regulation of ROS/PI3K/Akt signaling pathway [31]. The distant colonization of tumor cell metastasis is responsible for 90% of human cancer deaths [32]. C-X-C motif chemokine receptor 4 (CXCR4) is an important molecule for cancer migration and homing to the docking region. Rg3 treatment significantly inhibits the CXCR4 expression in MDA-MB-231 cells and reduces the number of migrating cells, causing chemotaxis and reducing the width of scars in wound healing [33]. Cancer stem cells retain self-renewal properties, which are responsible for cancer recurrence and resistance to anticancer therapies. Rg3 attenuated the expression of Sox-2 and Bmi-1 by inhibiting the nuclear localization of hypoxia-inducible factor-1α in MCF-7 mammary globules and blocked Akt-mediated self-renewal signaling to inhibit the specificity of breast cancer stem cells [34]. Mouse double minute 2 homolog (MDM2) oncogene is a negative regulator of p53 with a self-regulating loop between p53 and MDM2, which is amplified and overexpressed in various human cancers [35,36]. 25-OCH3-PPD suppresses cell migration and the expression of MDM2 and Epithelial–mesenchymal Transition (EMT) markers by inducing apoptosis and G1 blockade in vitro, leading to the inhibition of breast tumor growth xenografts in vivo [37]. Treatment of breast cancer MCF-7 cells with Rg5 stimulates cell apoptosis and cell cycle arrest at G0/G1 phase by up-regulation of p53, p21WAF1/CIP1, and p15INK4B expression, and down-regulation of cyclin D1, cyclin E2, and cyclin dependent kinases (CDK4) expression [38]. Our recent study showed that Rg5 induced apoptosis and autophagy by inhibition of PI3K/Akt signaling pathway against breast cancer in vivo, the tumor growth inhibition rate of high dose Rg5 (20 mg/kg) was 71.4 ± 9.4%, similar to the positive control docetaxel (72.0 ± 9.1%) [39]. The three major anti-breast cancer pathways of ginsenosides were summarized as shown in Figure 2.
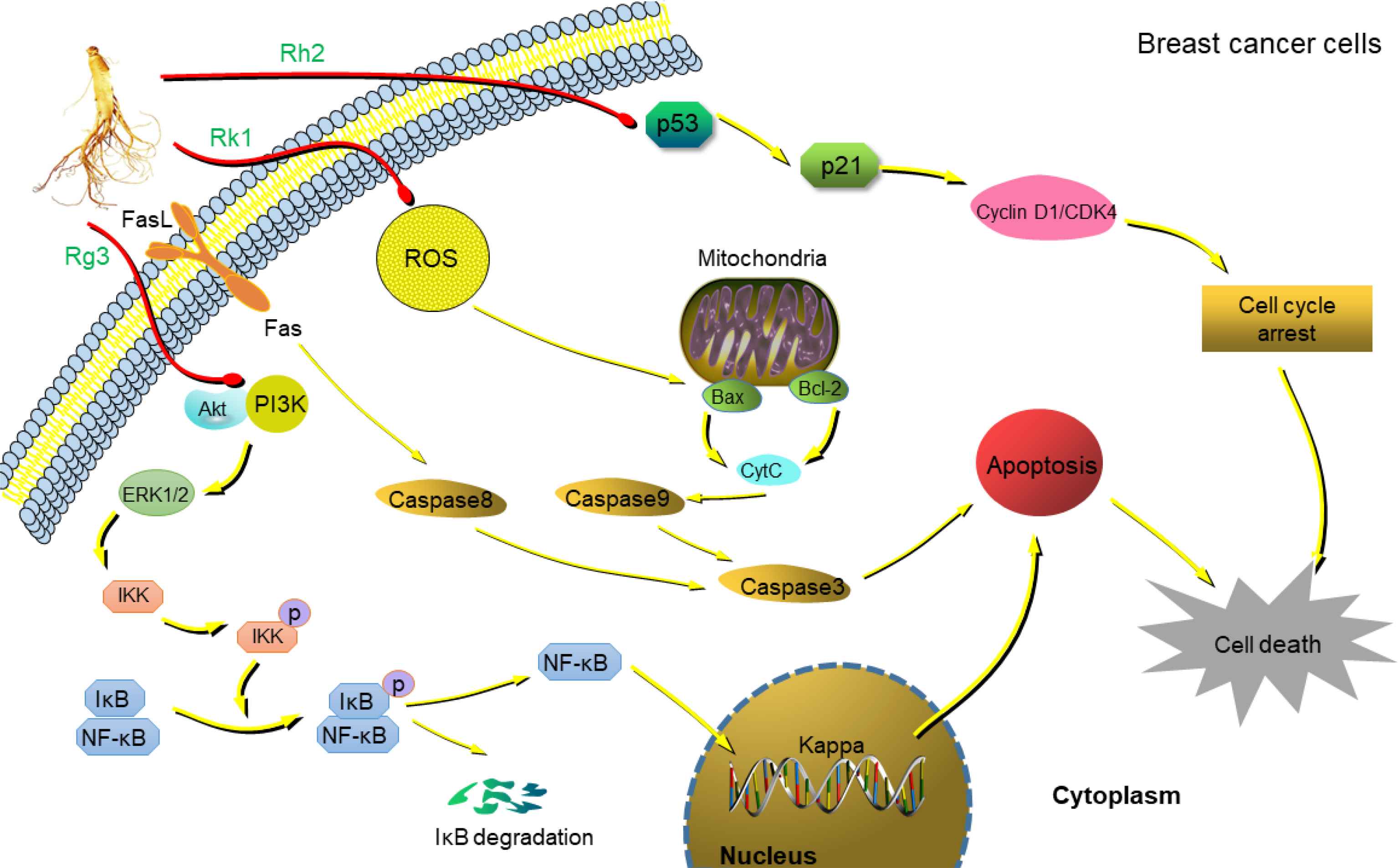
Anti-breast cancer mechanism of ginsenosides. Rh2 up-regulates the expression of Cyclin-dependent Kinase (Cdk) inhibitor p21WAF1/CIP1 by inducing the expression of cyclin p21, and induces cell cycle arrest in the G1 process [26]. Rg3 inhibits the phosphorylation of Akt and ERK, leading to inhibition of NF-κB causing apoptosis [30]. Rk1 triggers intracellular ROS production and reduces mitochondrial membrane potential, increases expression of Bax, cytochrome C, activates caspase 3, 8, and 9 levels, reduces Bcl-2 levels, blocks PI3K/Akt pathway, and induces cells apoptosis [31].
2.2. Colorectal Cancer
Colorectal Cancer (CRC) is a digestive system disease with high morbidity and mortality [40] and is the third most diagnosed cancer and the fourth most deadly cancer in the world [41]. It is reported that Rh2 stimulates apoptosis by activating the tumor suppressor factor p53 and inducing the proapoptotic regulator Bax in colorectal cancer cells. [42] The activation of transcriptional activator 3 (STAT3) is a major factor in the development of colon cancer, Interleukin-6 (IL-6) is a well-known and thoroughly studied cytokine in tumor-associated STAT3 signaling [43]. Rh2 effectively inhibits IL-6 induced signal transduction, STAT3 phosphorylation, and the expression of Matrix Metalloproteinases (MMPs), including MMP-1, -2, and -9, further prevents the invasion of cancer cells and enhances the sensitization of CRC cells to doxorubicin treatment [44]. Rg3 exhibits anti-angiogenic activity by reducing the incidence of peritoneal metastasis of intestinal adenocarcinoma induced by oxidized azomethane [45,46]. Rg3 is considered to be an effective adjuvant for the clinical treatment of colorectal cancer because it can inhibit the HCT116 cells growth by blocking the nuclear transport of β-catenin and decreasing the transcriptional activity of β-catenin/Tcf [46]. Additionally, combination of Rg3 and 5-fluorouracil down-regulates MMP levels to reduce the invasion of colon cancer SW480 cells, which may be because Rg3 enhances the cytotoxicity of 5-fluorouracil and oxaliplatin in xenografts [47]. The ginsenoside compound K (CK) is a gut microbial metabolite of Rb1 and a major component of American ginseng. CK exhibits significant anti-proliferative effects in HCT-116 and SW-480 cells at concentrations of 30–50 μM [48]. CK activates the expression of caspase-8 and -9 in HCT-116 and SW-480 cells and docking data indicates that CK forms hydrogen bonds with Lys253, Thr904, and Gly362 in caspase-8, as well as Thr62, Ser63, and Arg207 in caspase-9 arrested the progress of division and induces apoptosis [48]. Our group study found that Rh4 exhibits high anti-colorectal cancer activity with low toxicity and few side effects, and stimulates apoptosis and autophagic cell death via activation of the ROS/JNK/p53 pathway to inhibit the colorectal tumor growth [49]. EMT plays an important role in the migration, invasion, and metastasis of many types of cancer including CRC [50]. Rb2 binds to the hydrophobic pocket of Transforming Growth Factor (TGF-β1), which partially overlaps with the binding site on TGF-β1, thereby disrupting TGF-β1 dimerization and inhibiting the expression of Smad4 and Smad2/3 phosphorylation. Therefore, the TGF-β1/Smad/EMT signaling pathway may become a potential target for the treatment of colorectal cancer [51]. O-β-
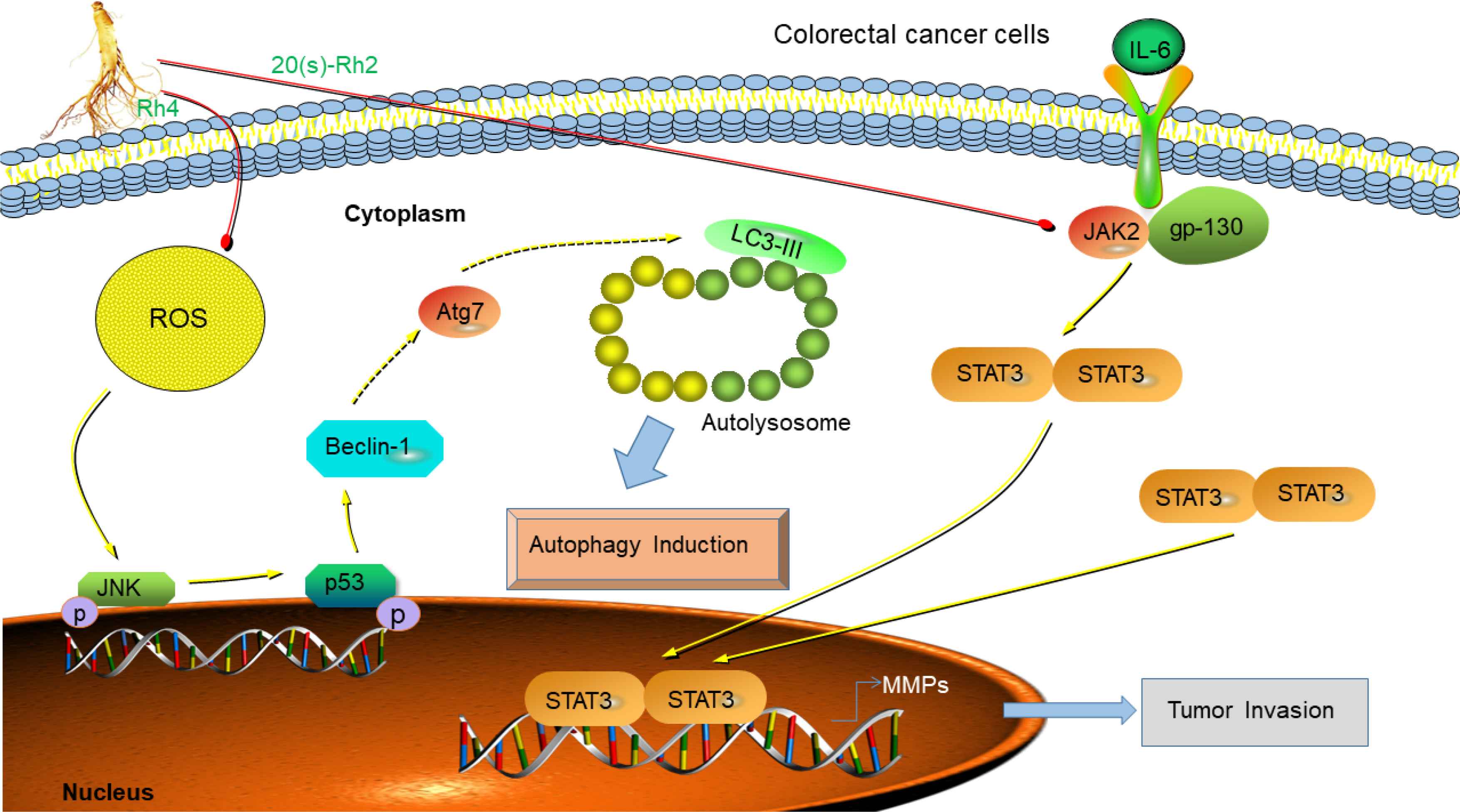
Anti-colorectal cancer mechanism of ginsenosides. 20(S)-Rh2 inhibits colorectal cancer by suppression of IL-6-induced signal transduction, STAT3 phosphorylation, and expression of MMPs, including MMP-1, -2, and -9 [44]. Rh4 induces colorectal cancer death by accumulation of ROS, activation of JNK-p53 signaling pathway and beclin-1 and Atg7 autophagy-related protein expression [49].
2.3. Ovarian Cancer
Ovarian cancer, cervical cancer, and breast cancer are known as the three major killers of female cancer [53]. According to 2018 statistics, ovarian cancer accounts for 4.4% of all female deaths from cancer [54], and the mortality is increasing year-by-year. Rg3 can significantly inhibit the metastasis of ovarian cancer [55,56], one potential reason is that Rg3 partially inhibits the tumor-induced angiogenesis, and decreases the invasive ability and MMP-9 expression of SKOV-3 cells [57]. 20(R)-Rg3 and 20(S)-Rg3 are stereoisomer formed by different orientations of OH groups on C-20, previous study indicated that 20(S)-Rg3 can inhibit the glycolysis of ovarian cancer cells by regulating Hexokinase 2 (HK2) and Pyruvate Kinase M2 (PKM2) by reducing the hypoxia-inducible factor-1α expression and activating the proteasome pathway [58]. 20(S)-Rg3 also inhibits EMT in ovarian cancer cells, thereby inhibiting cancer progression in vitro and in vivo via targeting the DNMT3A/miR-145/FSCN1 pathway [55,59], and inhibits the Warburg effect in ovarian cancer cells via regulation of H19/miR-324-5p/PKM2 pathway [56]. Rb1 is a potential therapeutic candidate for the treatment of ovarian cancer because it can reverse hypoxia-induced EMT by eliminating the inhibition of EP300 and E-cadherin miR-25 [60]. Cancer Stem Cells (CSCs) represents a subpopulation with self-renewal and differentiation capabilities that make these cells therapeutically tolerant. Rb1 and its metabolite CK specifically target the formation and expansion of CSCs, which can induce synergistic cytotoxicity by enhancing chemo-sensitivity to the clinical anticancer drugs cisplatin and paclitaxel, then inhibit cancer cell growth through Wnt/β-catenin signaling and EMT regulation [61,62]. Treatment of ovarian cancer SKOV3 cells with Rh2 significantly induces apoptosis by promoting the expression of lysed Poly-ADP Ribose Polymerase (PARP) and cleaved caspase-3, leading to inhibition of SKOV3 cells growth and its migrating invasive ability [63].
2.4. Gastric Cancer
Gastric cancer is the most commonly diagnosed digestive tract cancer with a high recurrence rate, high metastasis rate, and resistance to chemotherapeutic drugs [64]. Rg3 can induce apoptosis by activation of caspase-3, -8, and -9 in the human gastric cancer cell AGS by increasing the number of cells in the sub-G1 phase, caspase-3 activity, and the degree of PARP cleavage of the caspase-3 substrate [65]. The triol type triterpene glycoside Re can be converted into less polar ginsenosides, namely Rg2, Rg6, and F4, by heat treatment, which can activate the expression of caspase-3, -8, -9 and the cleaved PARP in AGS cells in a dose-dependent manner, further inhibits the phosphorylation of CDK2 at Thr160 by up-regulating p21 levels against the gastric cancer cells growth [66]. Helicobacter pylori CagA promotes the proliferation of gastric cancer cells through the overexpression of Fucosyltransferase (FUT4) in cells, tissues, and blood samples of gastric cancer patients [66]. Rg3 significantly induces FUT4-mediated apoptosis by activation of caspase-3, -8, and -9 and PARP expression in Helicobacter pylori CagA-treated gastric cancer cells through inhibition of FUT4 expression by up-regulation of Specific Protein 1 (SP1) and down-regulation of Heat Shock Factor protein 1 (HSF1) [67]. Rg3 can inhibit cell proliferation increase, migration, and invasion in TGF-β1 induced gastric cancer SNU-601 cell line by increased the expression of the epithelial marker E-cadherin, and repressed the expression of the mesenchymal marker Vimentin [68]. Rh2 can significantly induce apoptosis by up-regulation of Bax and down-regulation of Bcl-2 in a dose-dependent manner in human gastric cancer SGC7901 cells [69]. F2 treatment suppresses the SGC7901 growth by activation of p53 signaling pathway and the Bcl-xl/Beclin-1 pathway through up-regulation of some related protein expression such as Atg5, Atg7, Atg10, and PUMA, Bcl-xl, Beclin-1, UVRAG, and AMBRA-1, indicating that F2 can be used as a potential natural product for anti-gastric cancer [70].
2.5. Liver Cancer
Primary liver cancer is one of the most aggressive tumor types [71]. Early primary liver cancer is mainly treated by surgery (resection or transplantation), but the systemic metastasis of cancer and the prognosis of patients after relapse are poor [72]. Hepatocellular Carcinoma (HCC) is one of the most common forms of liver cancer and has a very poor prognosis. Sorafenib is the only drug approved by the U.S. Food and Drug Administration for the treatment of advanced HCC. However, this drug does not delay the progression of the symptoms of the disease and the monthly treatment cost is about 5400 US$ [73], also associated with serious side effects including the significant risk of bleeding [74]. More and more researches have indicated that natural product with low toxic side effects has become the focus for liver cancer treatment or prevention. Rho GTPase activating protein 9 (ARHGAP9) is closely related to tumor metastasis [75], inhibition of ARHGAP9 protein expression by Rg3 can significantly suppress the migration and invasion of human hepatoma cells HepG2 and MHCC-97L in vitro, and prevent the growth of HepG2 and MHCC-97L tumors in BABL/c nude mice in vivo [76]. Rg3 also can effectively inhibit cell proliferation and promote cell apoptosis through activation of endogenous mitochondria-mediated caspase-dependent apoptosis pathway in SMMC-7721 and HepG2 cells [77]. Previous studies have shown that Rh2 induced HepG2 cells apoptosis by activating the ROS-mediated lysosomal-mitochondrial apoptosis pathway [78]. The inhibitory effect of Rh2 on the migration ability of HepG2 cells is achieved by recruiting histone deacetylase, which leads to the inhibition of AP-1 transcription factors [79]. The treatment of HCC cells with Short Hairpin small interfering RNA (shRNA) that expresses Atg7 completely abolished the effect of Rh2 on β-catenin and cell viability, whereas β-catenin overexpression eliminated the effects of Rh2 on autophagy and cell viability [80]. In the canonical Wnt pathway, the binding of a Wnt protein ligand to a Frizzled family receptor activates β-catenin, whose nuclear translocation and retention lead to the regulation of gene transcription [81]. Rh2 increases autophagy and inhibits β-catenin signaling by inhibiting HCC cells growth [82]. In the Alcoholic Liver Disease (ALD) mice model, our recent study found that Rk3 significantly reduced AST and ALT levels in serum, reduced oxidative stress, restored the liver tissue antioxidant balance, and significantly reduced mice expression of inflammatory cytokines such as NF-κB, TNF-α, IL-6, and IL-1β [83]. The potential mechanisms of ginsenosides resistance to liver cancer are summarized in Figure 4.
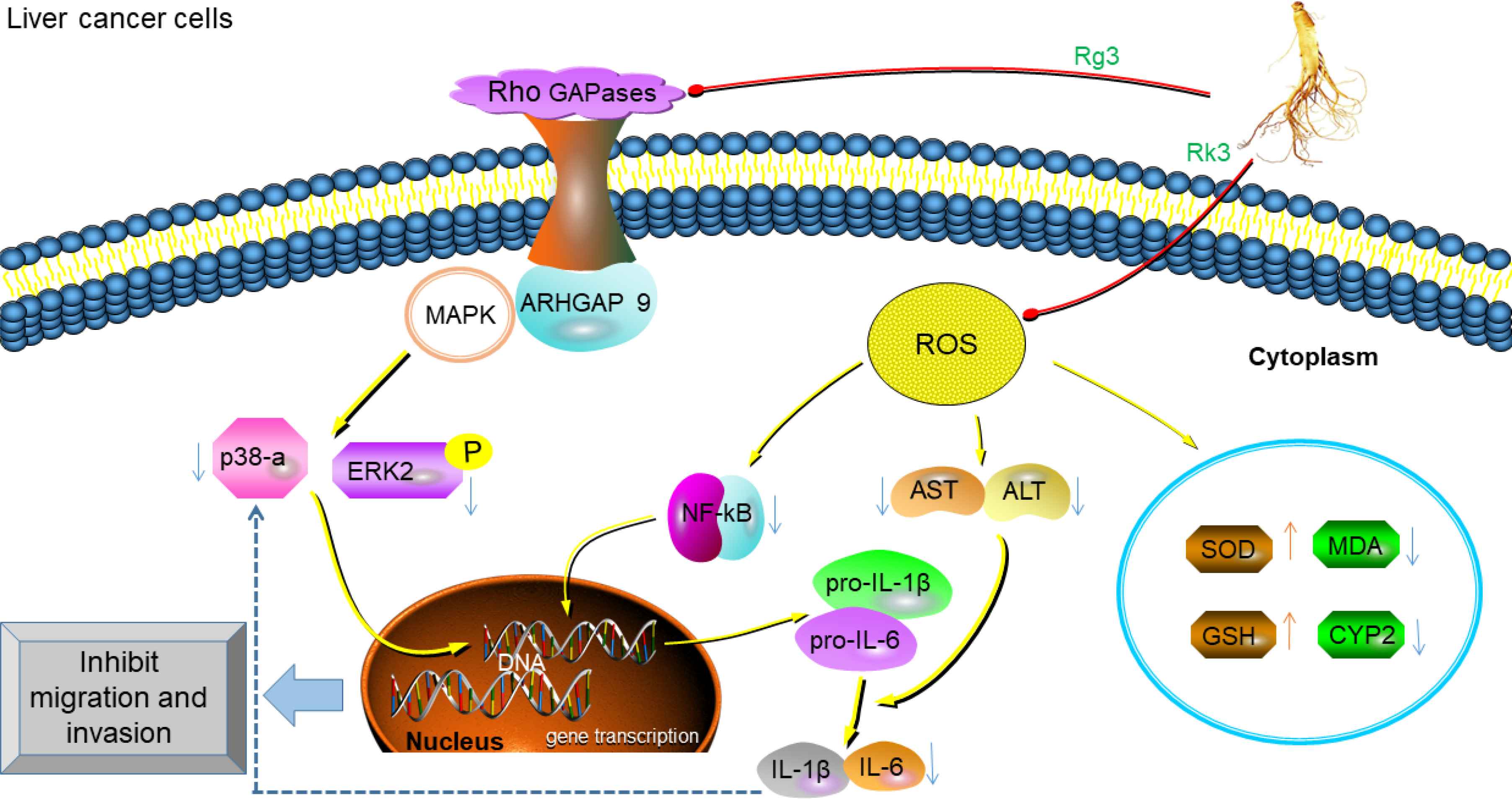
Anti-liver cancer mechanism of ginsenosides. Rg3 up-regulates the expression of ARHGAP9, and then in combination with MAPKs inhibits the activation of ERK2 and p38α in Swiss 3T3 fibroblasts, leading to the inhibition of the migration and invasion of liver cancer cells [76]. Rk3 suppresses NF-κB signaling pathway-mediated inflammatory cytokines expression to restore the antioxidant balance of liver tissue and promote liver function [83].
2.6. Lung Cancer
Lung cancer is the leading cause of cancer-related death worldwide and Lung Adenocarcinoma (LUAD). Despite the recent development of targeted therapies for some of the genetic subtypes of human LUAD, the overall survival rate of this cancer remains very low [84]. As mucosal tissue with the largest surface area in the body, the lungs inhale various airborne microbes and environmental contaminants. The development of lung cancer is closely related to chronic inflammation, which is characterized by the infiltration of inflammatory cells and the accumulation of pro-inflammatory factors, including cytokines, chemokines, and prostaglandins, that stimulate cell proliferation, angiogenesis, tissue remodeling, and metastasis [85]. The survival rate of PPT-treated SK-MES-1 cells is reduced to 66.8% and the therapeutic effect is close to that of cisplatin-treated lung cancer [86]. 20(R)-Rg3 epimer inhibits the EMT of lung adenocarcinoma by increasing the expression of E-cadherin and inhibiting the expression of vimentin and up-regulation of snail [87]. Rg3 also can inhibit the EMT and invasion of lung cancer by down-regulating FUT4-mediated EGFR inactivation and blocking the MAPK/NF-κB signaling pathway [88]. Rg3 can stimulate NF-κB expression by regulation of transcription factors including Cyclooxygenase-2 (COX2), MMP-9, VEGF, c-Myc, and cyclin D1 to make lung cancer tumors sensitive to radiation and stimulate apoptosis [89]. Rg3 can effectively inhibit the volume and weight of tumors in xenograft models and inducing apoptosis by inhibiting the PI3K/Akt signaling pathway [90]. Administration of Rg3 to lung cancer mice can improve the immune-regulation activities by increasing spleen cell proliferation [91]. Our previous research demonstrated that Rg3 treatment reduced the expression of cyclin D1 and CDK4 in Non-small Cell Lung Cancer Cells (NSCLC), up-regulated the expression of P21, inhibited cell proliferation and colony formation, and induced cell cycle arrest in G1 phase [89]. Rh2 prevents mitochondrial-dependent pathways by inducing ROS-mediated Endoplasmic Reticulum (ER) stress, thereby stimulating cisplatin resistance in NSCLC A549/DDP cells [92], it can also significantly induce apoptosis in A549 cells [93]. Mitomycin C (MMC) is an anticancer agent that causes intrachain DNA cross-linking and blocks DNA processes in transcription and replication levels [94]. It is reported that Total Ginsenoside extract (TGS) is a novel adjuvant anticancer compound, and combined treatment with TGS and MMC can effectively enhance the cytotoxicity of MMC against NSCLC in vitro and in vivo [95]. The potential mechanisms of anti-lung cancer are summarized in Figure 5.
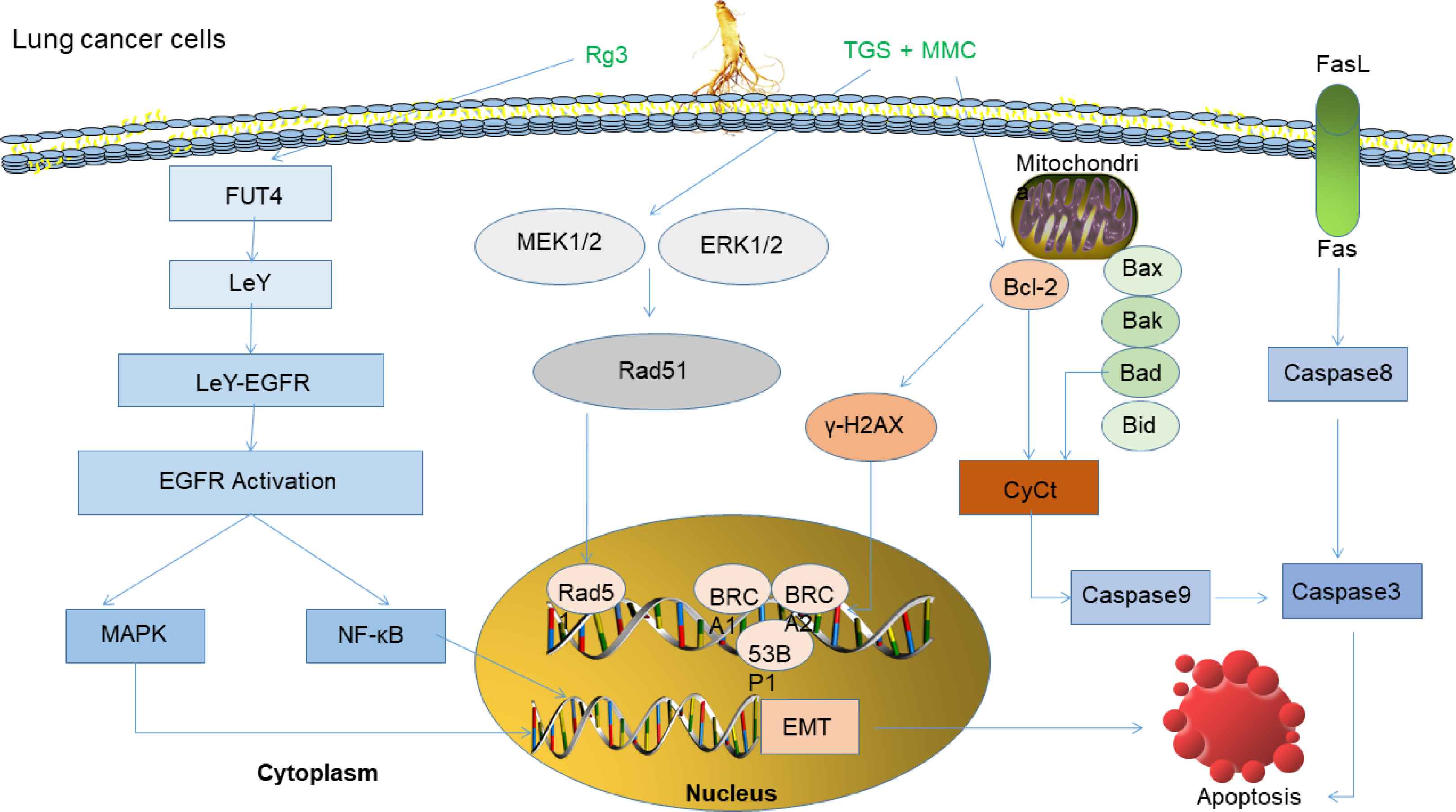
Anti-lung cancer mechanism of ginsenosides. Rg3 prevents FUT4 expression, LeY biosynthesis, and EGFR activation, then blocks MAPK and NF-κB signal pathways, leading to inhibition of migration, invasion and EMT of lung cancer cell [88]. In combination of TGS and MMC, the inhibitory effect of TGS on phosphorylated MEK1/2 and phosphorylated ERK1/2 significantly reversed MMC-induced S-phase cell cycle arrest and inhibited Rad51-mediated DNA damage repair [95].
2.7. Prostate Cancer
Prostate cancer is the second most commonly diagnosed cancer and the sixth leading cause of cancer death among men worldwide, with an estimated 1,276,000 new cancer cases and 359,000 deaths in 2018 [5]. Based on prostate-specific antigen screening and surgery for the diagnosis of latent cancer of benign prostatic hyperplasia [96], the diagnosis rate has decreased significantly in recent years, these declines are limited to early disease, while the incidence of late disease increases. At present, the clinical treatment of prostate cancer is radical local treatment plus/minus systemic treatment. Among patients receiving systemic treatment, the first choice is androgen deprivation therapy [97], but its side effects are large, so natural products need to be explored for treatment of prostate cancer. Rg3 (50 μM) combined with the conventional drug docetaxel (5 nM), or with cisplatin (10 μM) and doxorubicin (2 μM) inhibits prostate cancer cell growth and induces apoptosis by suppression of NF-κB activity and up-regulation of Apoptosis Inhibitory Protein (IAP-1), X chromosome IAP, and NF-κB targeted genes expression including Bax, caspase-3, and -9, and inhibited Bcl-2 [98]. Rg3 can effectively suppress migration of prostate cancer cells (PC-3M) by down-regulating AQP1 expression through activation of p38 MAPK pathway and some transcription factors acting on the AQP1 promoter [99]. The EC50s of Rg3 and Rh2 against the prostate cancer PC-3M cells were 8.4 and 5.5 μM, respectively, and those of LNCaP cells were 14.1 and 4.4 μM, respectively [100]. Both Rg3 and Rh2 induced apoptosis and regulated three MAP kinase activity modules in LNCaP and PC3M cells to inhibit the proliferation of prostate cancer cells [100]. PC-3M cells treated with processed ginseng (MG) showed the highest activity with a half-maximal inhibitory concentration of 48 μg/mL, which inhibited the growth of human prostate cancer cell xenografts in athymic nude mice [101].
2.8. Nasopharyngeal Carcinoma
Nasopharyngeal carcinoma (NPC) is a highly invasive and metastatic head and neck cancer. Distant metastasis is the main reason for the difficulty in the treatment of NPC. The common treatment for NPC is radiation therapy. Chemotherapy is another option for treating nasopharyngeal cancer patients, but resistance to conventional drugs is a challenge [102]. Rg3 can inhibit the expression of MMP-2 and -9 as well as EMT-related transcription factors, especially the zinc finger E-Box binding homeobox 1, which blocked cell migration and invasion in HNE1 and CNE2 cell lines [103], indicating that Rg3 may be a potentially effective agent for the treatment of NPC. Rg3 promotes lymphocyte proliferation in NPC patient with radiotherapy in a dose-dependent manner by up-regulating the ratio of CD4+/CD8+, the expression of IgG, IgM and IL-2, and down-regulating the expression of CD8+ and IL-6 [104]. Treatment with 20(S)-Rh2, CK, or PPD exhibits a significant role in inducing apoptosis of NPC cells (HK-1 cells) in a concentration-dependent manner [105]. CK most broadly inhibits HK-1 xenograft tumor growth and depletion of mitochondrial membrane potential depolarization induces AIF transfer from the cytoplasm of HK-1 cells to the nucleus [105]. Rh2 significantly inhibits the proliferation of nasopharyngeal carcinoma CSCs in vitro and promotes apoptosis [106]. Moreover, Rg1 can significantly inhibit the growth of anaplastic thyroid carcinoma and prolong the survival time of tumor-bearing mice [107].
2.9. Gallbladder Cancer
Gallbladder Carcinoma (GBC) is a malignant tumor of the biliary tract, accounting for 80–95% of the global biliary tract cancer and ranking sixth in gastrointestinal cancers with high mortality, late diagnosis and chemotherapy resistance [108]. Most patients were diagnosed with advanced GBC and cannot be treated by resection [109]. 20(R)-Rg3 acts as a dominant inhibitor of gallbladder cancer growth by activating the p53 pathway and subsequent cell senescence in vitro and in vivo [110].
2.10. Bladder Cancer
About 80% of bladder cancers are superficial tumors that can be easily treated with minimally invasive surgery. However, the recurrence of superficial transitional cell carcinoma of the bladder after transurethral resection is a major problem in the treatment of bladder cancer [111]. Since the late 1980s, cisplatin-based combination chemotherapy has become the standard treatment for advanced bladder cancer [112]. The serious side effects caused by anti-tumor drugs are the main obstacles to the success of their application and treatment, so the study of natural compounds with little or no toxicity has received much attention. Combination of Rg3 and cisplatin decreased cyclin Bcl-2 expression, but increased the expression of cytochrome C and apoptosis-related protein caspase-3 in T24R2 cells, indicating that the endogenous apoptotic pathway was activated [113].
2.11. Cervical Cancer
Cervical cancer is a major cause of cancer in women around the world with more than 510,000 new cases and 288,000 deaths each year [114]. Human Papillomavirus (HPV) infection is the most direct source of cervical cancer. HPV infection is usually asymptomatic and transient but can promote pre-cervical precancerous lesions. The treatment of cervical cancer, including radiation and chemotherapy, used together with various herbs [115], can significantly improve the treatment effects and increase the sensitivity of cancer cells. Rd inhibits the growth of human cervical cancer (HeLa) cells in a concentration and time-dependent manner, with an IC50 value of 150.5 ± 0.8 μg/mL incubation for 48 h. Its mechanism of action is to induce apoptosis by activating the caspase-3 pathway, reducing Bcl-2 expression and mitochondrial transmembrane potential and up-regulating Bax expression [116]. Sun Ginseng (SG), a major component of red ginseng, contains approximately the same amount of three major ginsenosides (RK1, Rg3, and Rg5) [117]. Combination of SG and epirubicin/paclitaxel synergistically induce the apoptosis of cervical cancer cells by increasing the caspase-9/-3 expression, the mitochondrial accumulation of Bax and Bak, and the cytochrome C release [117]. Rg5 exhibits significant genotoxic effects in HeLa and MS751 human cervical cancer cell lines [118]. 20(s)-Rg3 loaded magnetic human serum albumin nanospheres [20(S)-Rg3/HSAMNP] prepared by the desolvation-crosslinking method increased the water solubility and bioavailability of 20(S)-Rg3, combined with magnetic hyperthermia, it can effectively induce apoptosis of HeLa cervical cancer cells [119]. HeLa cells treated with microwave-irradiated product of ginseng (MG) can inhibit the growth of human cervical cancer xenografts in athymic nude mice in vivo, which is associated with cell death and the induction of autophagy [120].
2.12. Pancreatic Cancer
Pancreatic Cancer (PC) has a very high mortality rate with a 1-year survival rate of <10% [121]. The success rate of the surgical resection of PC is not high and the effect of chemotherapy is not work well. The current standard chemotherapy for pancreatic cancer is the pyrimidine analog Gemcitabine alone, or in combination with 5-Fluorouracil (5-FU) or a platinum reagent (e.g., cisplatin or oxaliplatin) [122]. However, the toxic side effects are accompanied by a certain degree of drug resistance, so combinations with natural products improve the drug sensitivity as an anticancer agent or adjuvant therapy. PC treatment with 25-OH-PPD and 25-OCH3-PPD can inhibit the MDM2 oncogenes and its related pathways to exert anticancer activity, which prevent the growth of xenograft tumors without any host toxicity [123]. Vasculogenic Mimicry (VM) is a novel tumor microcirculatory system that differs from classically described endothelium-dependent angiogenesis. VM is expressed in 71.79% (84/117) of the pancreatic cancer cases [124], so inhibition of VM channels by ginsenosides is extremely important to the prevention and treatment of PC. Rg3 effectively inhibits pancreatic cancer by down-regulating the expression of VE-cadherin, epithelial cell kinase (EphA2), MMP-2 and -9 [125]. As an adjuvant for the treatment of pancreatic cancer, Rg3 is used simultaneously with erlotinib to enhance the anti-proliferation and apoptosis effects in BxPC-3 and AsPC-1 pancreatic cancer cells and xenograft models by up-regulation of caspase-3, 9, and PARP cleavage expression and down-regulation of p-EGFR, p-PI3K, and p-Akt expression [126].
2.13. Melanoma
Melanoma is a type of skin cancer that develops from melanocytes located in the basal layer of the epidermis [127]. In the early stage, melanoma can be cured by surgical resection and the 5-year survival rate is >95% [128]. However, advanced metastatic melanoma is a malignant tumor. Due to drug resistance, there is no effective treatment. The involvement of MAPK [129], the PI3K/Akt pathway [130], and some mutant oncogenes, including BRAF [131], c-KIT [132], provides new potential targets for the diagnosis and treatment of melanoma. Rg3 has been shown to reduce melanoma cell proliferation by reducing histone deacetylase 3 and increasing p53 acetylation [133]. Rg3 also can reduce the expression of ERK and Akt in vivo and in vitro, thereby down-regulating VEGF in B16 cells, weakening the proliferation and migration of vascular endothelial cells, and inhibiting blood vessels generate [134].
To facilitate the readers to understand the antitumor activity of different types of ginsenosides on different types of cancer in recent years, we have classified the main information in Table 2.
| Cancer type | Cells/Animal model/Human clinical trials | Ginsenosides | Doses | Durations | Biological effects | References |
|---|---|---|---|---|---|---|
| Breast cancer | MDA-MB-231 cell xenograft nude mice | Rk1 | 10–20 mg/kg/day | 21 days | Inhibited cell proliferation and repressed tumor growth by ROS/PI3K/Akt pathway | [31] |
| MDA-MB-231 cells | GER | 1–2.5 mg/mL | 48 h | Inhibit the self-renewal ability of stem cell-like breast cancer cells | [34] | |
| MDA-MB-468 cell xenograft nude mice | 25-OCH3-PPD | 20 mg/kg/day | 4 weeks | Down-regulation EMT/MDM2 to induction of apoptosis and G1 phase arrest | [37] | |
| BALB/c nude mouse | Rg5 | 10–20 mg/kg/day | 30 days | Induction of apoptosis and autophagy | [39] | |
| MCF-7 cells | Rh2 | 20–50 μM | 24 h | Induces expression of cyclin p21 and decreases cyclin D3 levels, enhances immunogenicity and inhibits cancer cell growth | [26] | |
| MDA-MB-231 cells | Rg3 | 0–30 μM | 24 h | Promote apoptosis by ROS/PI3K/Akt pathway | [30] | |
| Colorectal cancer | Wistar rats | Rg3 | 2.5–5 mg/kg/3 days | 6 weeks | Inhibit peritoneal metastasis of intestinal adenocarcinoma | [46] |
| Caco-2 cells xenograft nude mice | Rh2 | 20–40 mg/kg/day | 30 days | Induction of apoptosis and autophagy by ROS/JNK/p53 pathway | [49] | |
| HCT-116 and SW-480 cells | CK | 30–50 μM | 48 h | Anti-proliferative effects | [48] | |
| Ovarian cancer | SKOV-3 cell tumor-bearing mice | Rg3 | 053 mg/kg/day | 20 days | Inhibits ovarian tumor-induced angiogenesis and lung metastasis | [55] |
| SKOV-3 cells | 20(S)-Rg3 | 80 μg/mL | 24 h | Inhibit cancer cell invasion and metastasis | [57] | |
| 3AO cells | 160 μg/mL | |||||
| SKOV-3 cells | Rb1 | 0–320 μg/mL | 24 h | Inhibit cancer metastasis by EMT | [60] | |
| Prostate cancer | PC-3 cells | Rg3 | 10 μM | 24 h | Down-regulation of AQP1 expression through the p38 MAPK pathway effectively inhibits migration of PC-3M cells | [99] |
| PC-3 cells | Rh2 | 150–200 μM | 24 h | Inhibit proliferation | [100] | |
| LNCaP cells | ||||||
| DU145 cells | MG | 50–100 μg/mL | 24 h | Induction of apoptosis and autophagy | [101] | |
| LNCaP cells | ||||||
| PC-3 cells | ||||||
| Athymic xenograft nude mice | 200 mg/kg/day | 5 weeks | ||||
| Gastric cancer | AGS cells | 20(S)-Rg3 | 0–100 μg/mL | 24 h | Induction of caspase-3, -8 and -9 activation leading to apoptotic cell death | [65] |
| SGC7901 cells | F2 | 20 μM | 12 h | Activation of ribosomal protein-p53 signaling pathway and Bcl-xl/Beclin-1 pathway inhibit cancer cells | [70] | |
| Lung cancer | A549 | Rg3 | 0–100 μg/mL | 24–48 h | Down-regulates FUT4-mediated EGFR inactivation and blocks MAPK and NF-κB signaling pathways to inhibit EMT and lung cancer invasion | [88] |
| H1299 | ||||||
| H358 | ||||||
| A549 cell xenograft nude mice | 10 mg/kg/3 days | 3 weeks | ||||
| H460 cell xenograft nude mice | Rg3-RGP | 100 mg/kg/day | 28 days | Antitumor activities via indirect immunomodulatory actions | [91] | |
| A549 cells | Rh2 | 25 mg/L | 48 h | Inhibit cell proliferation | [93] | |
| A549 cells | TGS + MMC | 1 mg/mL | 24 h | Reverses MMC-induced S-phase cell cycle arrest and inhibits Rad51-mediated DNA damage repair, enhancing cytotoxicity to cancer cells | [95] | |
| Liver cancer | HepG2 cells | Rg3 | 0–5 μg/mL | 24 h | Suppressed the migration and invasion of liver cancer cells by up-regulating the protein expression of ARHGAP9 | [76] |
| MHCC-97L cells | ||||||
| BABL/c nude mice | 0–10 mg/kg/day | 21 days | ||||
| HepG2 cells | Rh2 | 80 μM | 24–72 h | Reduce the expression levels of MMP3 gene and protein inhibit migratory ability | [79] | |
| BABL/c nude mice | Rk3 | 25–50 mg/kg/day | 6 weeks | Antioxidant, anti-apoptotic, and anti-inflammatory activities | [83] | |
| Nasopharyngeal carcinoma | HNE1 and CNE2 cells | Rg3 | 100 μg/mL | 24 h | Inhibits migration and invasion by regulating the expression of MMP-2 and MMP-9 and inhibiting EMT | [103] |
| HK-1 cells | CK | 15 μM | 24 h | Apoptosis mediated through the mitochondrial pathway | [105] | |
| CNE-2S cells | Rh2 | 30–60 μM | 6 days | Apoptosis | [106] | |
| Gallbladder cancer | NOZ cell xenograft nude mice | 20(S)-Rg3 | 20–40 mg/kg/day | 3 weeks | Activation of the p53 pathway and subsequent induction of cell senescence and mitochondrial-dependent apoptosis to attenuate GBC growth | [110] |
| NOZ GBC-SD cells | 0–400 μM | 48 h | ||||
| Bladder cancer | T24R2 cells | Rg3 | 50 μM | 48 h | Activation of the intrinsic apoptotic pathway and the enhancement of cell cycle alterations | [113] |
| Cervical cancer | HeLa cells | SG | 80 μg/mL | 48 h | Significantly enhanced the anticancer activity of epirubicin and paclitaxel in a synergistic manner | [117] |
| HeLa cells | Rg5 | 5 μM | 48 h | Induces apoptosis and DNA damage | [118] | |
| MS751 cells | 10 μM | |||||
| Pancreatic | PANC-1 cells | Rg3 | 10–80 μM | 24 h | Induction of apoptosis and down-regulation of the EGF/PI3K/AKT pathway to enhance the efficacy of erlotinib in inhibiting the proliferation of pancreatic cancer cells | [126] |
| Melanoma | B16 cells | Rg3 | 0–5 μg/mL | 48 h | Mediated through suppression of ERK and Akt signaling to inhibit tumor growth | [134] |
The inhibitory effect of different types of ginsenosides on different types of cancer
3. CONCLUSION
Ginsenosides play a role in the complex process of tumor development, including proliferation, apoptosis, migration, angiogenesis, and tumor immunogenicity [135]. Unlike targeted drugs, ginsenosides have multiple targets and exhibit anticancer activity through a variety of mechanisms, including targeting multiple tumor-associated signaling pathways and regulating intracellular ROS [136]. For example, the inhibition of proliferation (cyclin D1, COX-2, angiogenic factors (interleukin-8 and VEGF), invasion (MMP-9 and NF-κB) activation and metastasis and its down-regulation of gene products regulated by NF-κB, which have a significant role in anti-apoptosis [Cytostatics of IAP-1 (cIAP-1), Bcl-2, apolipoprotein precipitation protein inhibitors and Bcl-xl]. By referring to a large number of literatures, ginsenosides, through activation of the p53 tumor suppressor gene, cooperate with other cytokines to inhibit tumor growth, metastasis and promote apoptosis are the most common mechanisms for ginseng to treat different cancers (Figure 6). Due to the high toxicity of anticancer drugs and serious damage to organs, and with the gradual clarification of cancer pathogenesis, the exploration of ginsenosides as effective anticancer drugs or anticancer adjuvants holds good prospect.
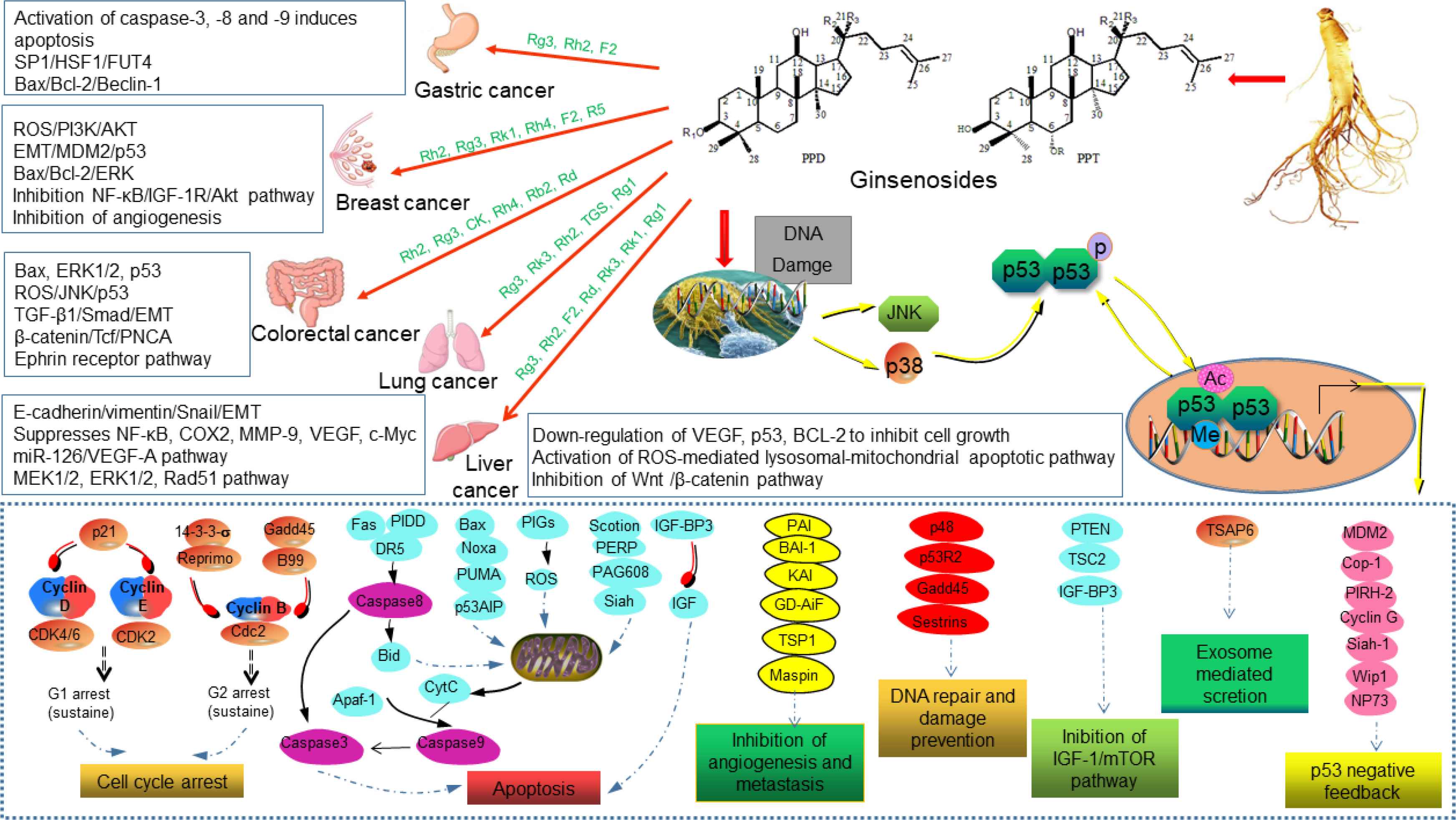
Activation of the p53 tumor suppressor gene plays a key role in cancer therapy. The action of ginsenosides on different targets allows p53 to be ubiquitinated, phosphorylated, interacts with transcriptional cofactors, and ultimately is important for activating target genes and responses (e.g., cell cycle arrest, DNA repair, apoptosis, and senescence) [35,38]. The non-receptor tyrosine kinase c-Abl can also be activated by DNA damage, then activates JNK/p38 and leads to p53 activation, which has a significant inhibitory effect on cancer cells [76].
CONFLICTS OF INTEREST
The authors declare they have no conflicts of interest.
AUTHORS’ CONTRIBUTION
HC and JD contributed to study design and writing original draft preparation. HY and DF contributed to revising and editing. All authors read and approved the final manuscript.
ACKNOWLEDGMENTS
This work was financially supported by the
Footnotes
REFERENCES
Cite this article
TY - JOUR AU - Hongwei Chen AU - Haixia Yang AU - Daidi Fan AU - Jianjun Deng PY - 2020 DA - 2020/05/22 TI - The Anticancer Activity and Mechanisms of Ginsenosides: An Updated Review JO - eFood SP - 226 EP - 241 VL - 1 IS - 3 SN - 2666-3066 UR - https://doi.org/10.2991/efood.k.200512.001 DO - 10.2991/efood.k.200512.001 ID - Chen2020 ER -