
The decrease of Tie-2 receptor phosphorylation in microvascular endothelial cells is involved in early brain injury after subarachnoid hemorrhage
- DOI
- 10.1016/j.artres.2018.07.004How to use a DOI?
- Keywords
- Tie-2 receptor; Microvascular endothelial cells; Early brain injury; Subarachnoid hemorrhage; Tight junction
- Abstract
Purpose: Loose endothelial cells and the destruction of the blood–brain barrier (BBB) are one of the pathophysiological mechanisms of early brain injury (EBI) after subarachnoid hemorrhage (SAH). Tie-2 receptor phosphorylation is important for maintaining integrity of microvascular endothelial cells and BBB. This study aimed to explore the role and changes of Tie-2 receptor phosphorylation levels in EBI after SAH.
Methods: This study used an endovascular puncture model of rat to simulate the occurrence and development of aneurysmal subarachnoid hemorrhage. The location of Tie-2 receptor in brain tissues was determined by immunofluorescence. Immunofluorescence and western blot was carried out to observe the expression of Claudin-5 and Occludin in cortex and hippocampus. We chose to observe the Tie-2 receptor phosphorylation level in hippocampus according western blot. Evans blue viability assay was used to evaluate BBB permeability.
Results: The results suggested that Tie-2 receptor mainly expressed around the vascular endothelial cells in brain. Following SAH, the Tie-2 receptor phosphorylation level and expression of tight junction protein (claudin-5 and occluding) decreased. Both of these downtrends were reversed by exogenous Angiopoietin-1 (Ang-1). Finally, injection of exogenous Ang-1 reduced SAH-associated BBB leakage.
Conclusions: Our study indicated that Tie-2 receptor phosphorylation in microvascular endothelial cells was involved in pathophysiological process after SAH, and the decline of Tie-2 receptor phosphorylation might increase blood–brain barrier permeability by decreasing the tight junction protein expression.
- Copyright
- © 2018 Association for Research into Arterial Structure and Physiology. Published by Elsevier B.V. All rights reserved.
- Open Access
- This is an open access article distributed under the CC BY-NC license.
Introduction
Subarachnoid hemorrhage (SAH) is a common cause of acute cerebrovascular disease, including intracranial aneurysm rupture of spontaneous SAH etiology of about 85%.1 According to epidemiological statistics, the overall incidence of spontaneous SAH is 10 people/10 million per year, ranking third in cerebrovascular disease, and second only to Cerebral thrombosis and cerebral hemorrhage.2 The mortality rate of aneurysmal subarachnoid hemorrhage reaches up to 51%, and 46% of survivors remain significant neurological dysfunction.3,4
The blood brain barrier (BBB) is a special barrier system in the central nervous system. It plays an important role in selectively absorbing nutrients, blocking pathogens and maintaining the central nervous system steady state.5 One of the earliest signs of brain injury after SAH is the change in BBB permeability, including microvascular endothelial cell apoptosis, opening of cell–cell tight junctions, and degradation of basal membrane extracellular matrix (ECM) and type IV collagen.6,7 All of the above pathophysiological process increase the brain edema, microthrombosis, inflammatory response and abnormal brain metabolism, and result in irreversible damage.
Brain microvascular endothelial cells (BMECs) are the important components of BBB, and tight junctions between BMECs are the key factors in ensuring the physical barrier integrity of BBB. A recent animal experiment found that tight connection loosening between the BMECs occurred in the hyperacute period (3h) after SAH, and resulted in BBB leakage and angiogenic brain edema.8 Angiopoietin (Ang) is an important endothelium-specific angiogenic factor. Unlike vascular endothelial growth factor (VEGF), the Angiopoietin/Tie-2 system mainly acts on the later stages of angiogenesis, inducing and sustaining endothelial cell migration and survival, and plays an important role in vascular stabilization.9 Angiopoietin-1 (Ang-1) and Angiopoietin-2 (Ang-2) are the most important proteins in the currently known angiopoietin family. Their binding targets are Tie-2 receptors on the surface of vascular endothelial cells. Ang-1 can make Tie-2 receptor phosphorylation, while Ang-2 cannot. The TIE-2 receptors which bind to ANG-1 promote the maturation and stabilization of endothelial cells, inhibit the inflammatory response and increase the cell–cell tight junction.10 Ang-2 binds to the Tie-2 receptor and competitively inhibits the effect of Ang-1, allowing endothelial cells to loosen, migrate, promote inflammatory responses, and disrupt cell connections.11 Under physiological conditions, Ang-1 expression in the brain tissue is higher, and Ang-2 is stored in the Weibel-Palade bodies of endothelial cells. So the highly phosphorylated Tie-2 receptors of BMECs ensure the endothelium Cell stability.
In 2011, Fischer et al.12 found that the serum concentration of Ang-1 was significantly changed within 1 day in a single-center clinical study of patients with subarachnoid hemorrhage. This finding was confirmed by Yi Wang et al., in 2015,13 who found that patients with higher Ang-1 serum concentrations had better prognosis in another larger sample size study. Furthermore, our published basic study has demonstrated that, in the early period after SAH, Ang-1/Ang-2 ratio decreased significantly in rat brain tissue.14
These all encouraging studies suggest that Angiopoietin/Tie-2 signal path is involved in the pathophysiology of subarachnoid hemorrhage. So it is necessary to explore the mechanism. On this basis, we suspect that Angiopoietin/Tie-2 signal path plays a role in BBB permeability via influencing the stability of microvascular endothelial cells after subarachnoid hemorrhage.
Methods
Animals and experimental tissue acquisition
120 adult male Sprague–Dawley rats weighing 250–300 g (Experimental Animal Center of Huzhou Teachers College) was housed in 12hr light and dark cycles in a temperature and humidity controlled environment.
In the first part, euthanasia was carried out in the Sham-operated group (n = 10) at 24h, while the SAH group at 6h (n = 10) and 24h (n = 10). We washed the brains from the rest in normal saline solution and fixed them overnight at room temperature in 4% paraformaldehyde in 0.01 M phosphate buffered saline. We then dehydrated the tissue blocks with an ascending ethanol solution series, cleared with xylene, and then embedded in paraffin. We cut the paraffin blocks into transverse serial sections of 10-μm thickness. Next, from each perfused rat, we randomly took five sections containing the hippocampus and cortex and mounted them on poly-
In the second part, the remaining rats were divided into Sham group (n = 30), SAH group (n = 30) and SAH + Ang-1 group (n = 30). After euthanized with perfused with saline at 24h, two thirds of rats were used for Western Blot (10 per group) and immunofluorescence staining (10 per group). The rest were used for Evans Blue viability assay.
This study followed strict accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health. The protocol was approved by the Biomedical Ethics Committee of Medical College of Huzhou Teachers College.
The model of SAH
After anesthetized using an intraperitoneal injection of ketamine-xylazine (80 mg/kg), experimental rats of SAH model was established using a standard endovascular puncture protocol. We introduced a sharpened prolene 3.0 suture into the right external carotid artery and advanced it through the internal carotid artery, then closured the common carotid artery near the junction of external and internal carotid arteries. We further advanced the suture into the intracranial internal carotid artery until we felt the resistance and then pushed 3 mm further into the internal carotid artery near the bifurcation with the middle cerebral artery. The duration of endovascular occlusion was between 30 s and 2 min. The carotid artery occlusion was necessary in the surgical operation for avoiding intraoperative bleeding. But excessive occlusion can lead to cerebral infarction, so we limited the occlusion time to within 2 min. This procedure was strictly enforced in all rats. Sham-operated rats underwent exactly the same surgical procedure, however, without puncturing the bifurcation of the internal carotid artery.
Drug administration
After inducing SAH we injected the rats with Ang-1 peptide (Abcam, ab109637) at 0 min. According to previous research results,14 we chose 50 μg as a therapeutic dose. So we injected 50 μg peptide through the tail vein in about 3 min. We euthanized half of the rats from second set of experiments and used the brains for Evans Blue viability assay and Western Blot. The other half animals were subjected to immunofluorescence.
Immunofluorescence
Immunofluorescence was carried out to locate Tie-2 receptor and to observe the expression of Claudin-5 and Occludin in rat brain. Sections were firstly deparaffinized and rehydrated. Then sections were incubated with 0.2% triton-100 at 23 °C for 15 min. In the next step, sections were soaked in citrate buffer (10 mmol/L, pH 6.0), and heated at 90 °C for 30 min, cooled at 23 °C, and then rinsed in PBS. We blocked Non-specific protein binding by 1 h incubation in 5% goat serum. Samples were incubated with primary antibodies (monoclonal mouse anti Tie-2 receptor anti-body, 1:100, Abcam; monoclonal mouse anti Claudin-5 antibody, 1:50, SantaCruz; monoclonal mouse anti Claudin-5 antibody, 1:50, SantaCruz) overnight at 4 °C, followed by a 15 min rinsing in PBS. Samples were then incubated with fluorescently labeled secondary antibodies (goat anti-mouse FITC A22110 IgG, 1:100, Abbkine) for 60 min at 23 °C, followed by a 15 min rinsing. Afterwards, samples were incubated with DAPI for 15 min at 23 °C. The pictures were collected from fluorescence microscopy. Samples without saline perfusion were taken as the positive control, while with PBS instead of primary antibodies as the negative control.
Western blot analysis
Using a tissue homogenizer we solubilized the frozen tissue samples in RIPA buffer on ice. We separated 60 μg/lane of protein samples on a sodium dodecyl sulfate (SDS)–polyacrylamide gel (10 or 12% acrylamide gel) and electrotransferred onto polyvinylidene fluoride (PVDF) membranes. After 2h incubation in a blocking solution (5% non-fat milk in 20 mM Tris–HCl, 150 mM NaCl, 0.1% Tween-20; TBST), we blotted the membranes with primary antibodies against pTie-2 (1:1000, Abcam, ab192800), Tie-2 (1:1000, Abcam, ab24859), Claudin-5 (1:100, SantaCruz, sc374221), Occludin (1:100, SantaCruz, sc133256), NADPH (1:1,000, Beijing Biosynthesis), or b-ctin (1:1,000, Beijing Biosynthesis) overnight at 4 °C. After extensive rinsing with TBST buffer, we incubated the membranes with HRP-conjugated secondary antibodies. We detected the antigen specific bands using chemiluminescence kit (provider) and X-ray films (provider). We performed western blots on at least six independent experiments. After scanning the X-ray films, we analyzed the density of the bands with Quantity one software version 4.62 (Bio-Rad, USA).
Evans Blue viability assay
24h after the models were established successfully, we randomly selected 10 rats from each experimental group of the second set of experiments for Evans blue viability assay. Briefly, we anesthetized the rats with an intraperitoneal injection of 10% chloral hydrate (0.35 ml/kg) and infused with 37 °C Evans blue dye (2% in 0.9% normal saline, 4 ml/kg) via the right femoral vein, injected over 5 min. Two hours later, we perfused the rats with 300 ml saline to wash out any remaining dye in the blood vessels. We then removed brains and cut them into two mm thick sections. The brain slices were weighed and immersed into formamide. We spectrophotometrically evaluated the BBB permeability in the brain motor cortex tissue by measuring the absorbance of Evans blue in the supernatant (Molecular Devices OptiMax, USA) at 610 nm.
Statistical analysis
For statistical analysis we used SPSS 19.0 software (SPSS Inc., Chicago, IL, USA). We present the data as mean ± SD. We used ANOVA to compare multiple groups. The LSD tests were used for to compare the differences between the two groups. Either P < 0.05 or P < 0.01 was considered statistically significant.
Results
General observations
In this study, the mortality is 0% in normal and sham groups, while totally 14 (17.5%) rats from SAH related groups died. Dead rats were excluded from further analysis. Every brain of SAH group was photographed immediately after removal to determine the success of the model. We also observed the expected decrease in post-surgical physical activity.
Location of Tie-2 receptor in rat brain after SAH
We used immunofluorescence to show the location of Tie-2 receptor in hippocampus. In the sham group, Tie-2 receptor was found gathered near the hippocampus vascular area (Fig. 1). However, Tie-2 receptor showed a lower expression in neuronal dense areas. Similar results was found in SAH 6h and SAH 24h groups. It indicated that the Tie-2 receptor was mainly expressed around the vascular endothelial cells.
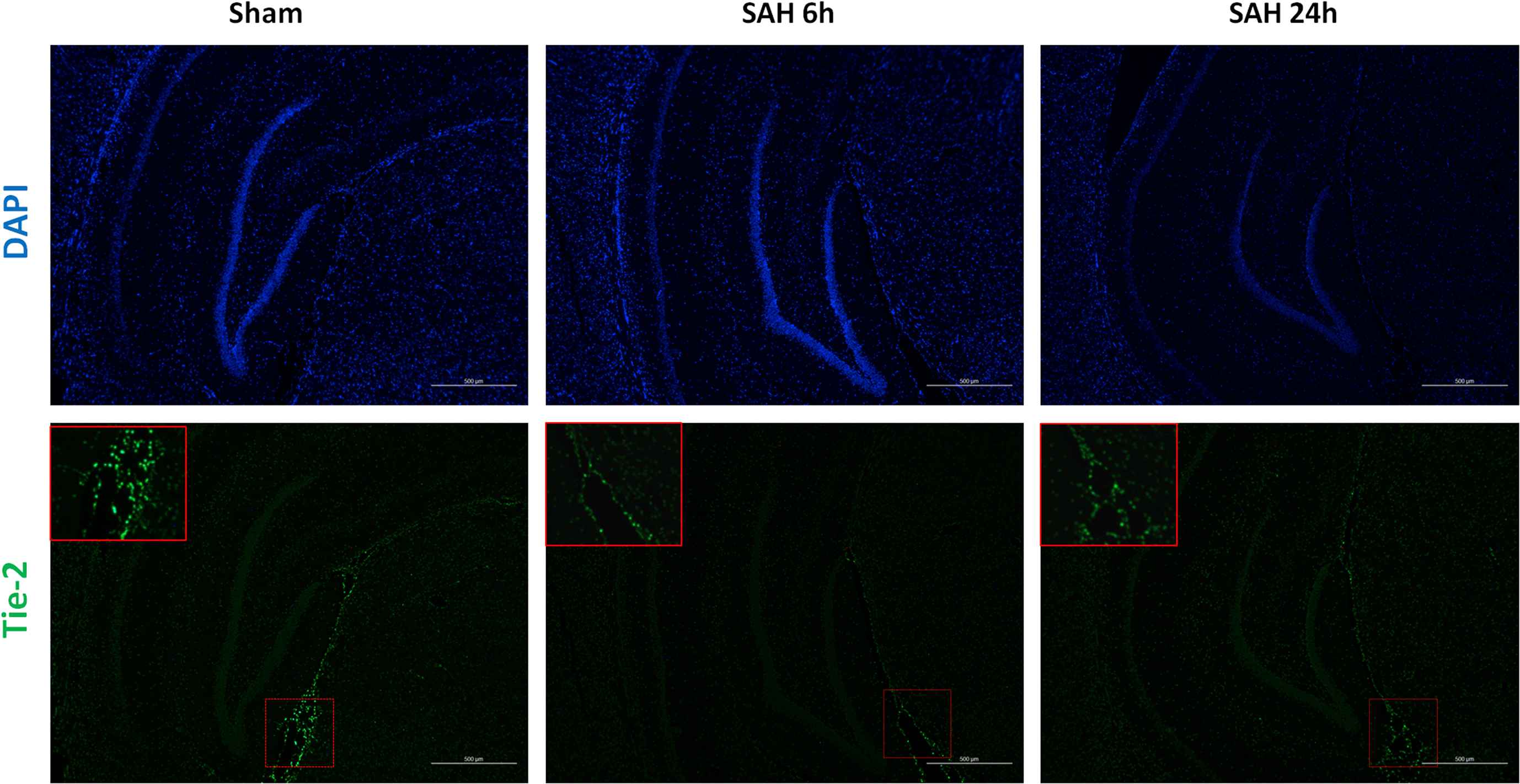
Immunofluorescence (×200) of Tie-2 receptor. All cell nucleuses were stained blue with DAPI, while Tie-2 receptor was stained green (shown and magnified in red frame). (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
Tie-2 receptor phosphorylation in hippocampus by western blot
In the first part of the experiment, we have found that Tie-2 receptor was mainly expressed in the dense area of blood vessels in hippocampus. The main function of the tie-2 receptor is initiated by phosphorylation. In the second phase of the experiment, we chose to observe the Tie-2 receptor phosphorylation level in hippocampus according western blot. The results demonstrated that, in sham-operated group, Tie-2 receptor maintained high level of phosphorylation, which decreased significantly after SAH (Fig. 2). The level of Tie-2 receptor phosphorylation, previously reduced, was improved after Ang-1 injection.
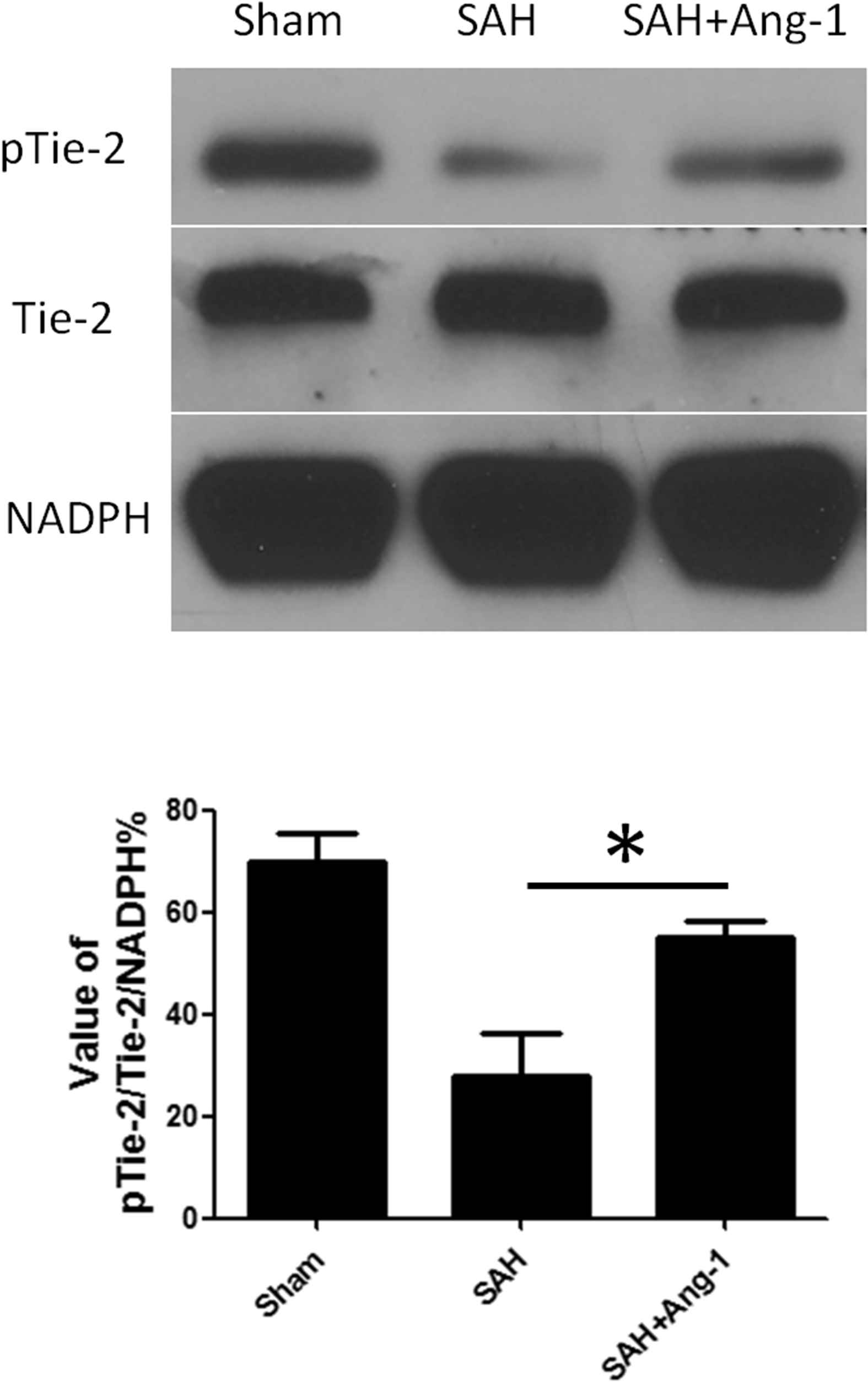
Western blot and quantitative analysis of Tie-2 receptor phosphorylation in hippocampus. Results are described as mean ± standard deviation. Ang-1, angiopoietin-1; SAH, subarachnoid hemorrhage. *P < 0.05. versus the SAH group.
Impact of exogenous Ang-1 on tight junction protein expression
We used both of immunofluorescence and western blot to show the expression of tight junction protein, Claudin-5 and Occludin, in cortex and hippocampus. Claudin-5 and Occludin were located around the endothelial cells of pia vessels in cortex by immunofluorescence (Fig. 3a). In hippocampus, tight junction protein mainly expressed around the deep capillaries (Fig. 3b). The distinct decrease in Claudin-5 expression from SAH group was observed via immunofluorescence. The results of western blot from hippocampus showed the same trend (Fig. 4). Similar results were seen in Occludin expression.
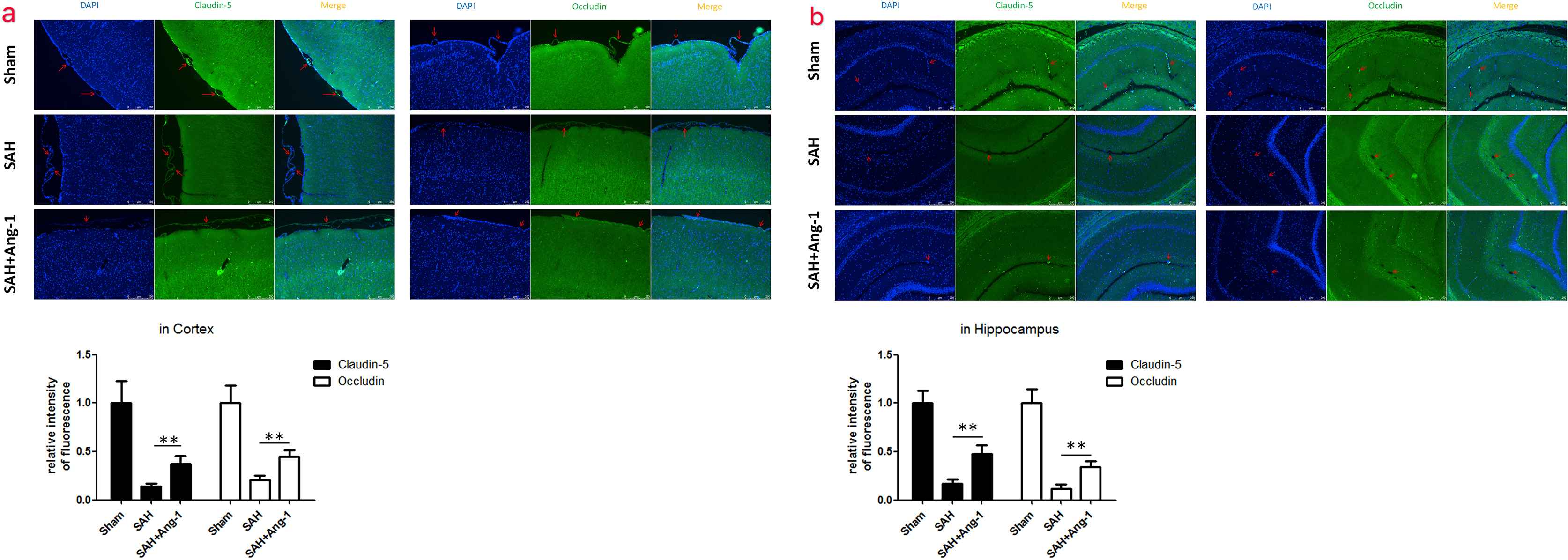
Immunofluorescence (×200) of Claudin-5 and Occludin in cortex (a) and hippocampus (b). All cell nucleuses were stained blue with DAPI, while Claudin-5 or Occludin was stained green. The red arrow showed the microvessels. Results are described as mean ± standard deviation. Ang-1, angiopoietin-1; SAH, subarachnoid hemorrhage. **P < 0.01. versus the SAH group. (For interpretation of the references to color in this figure legend, the reader is referred to the Web version of this article.)
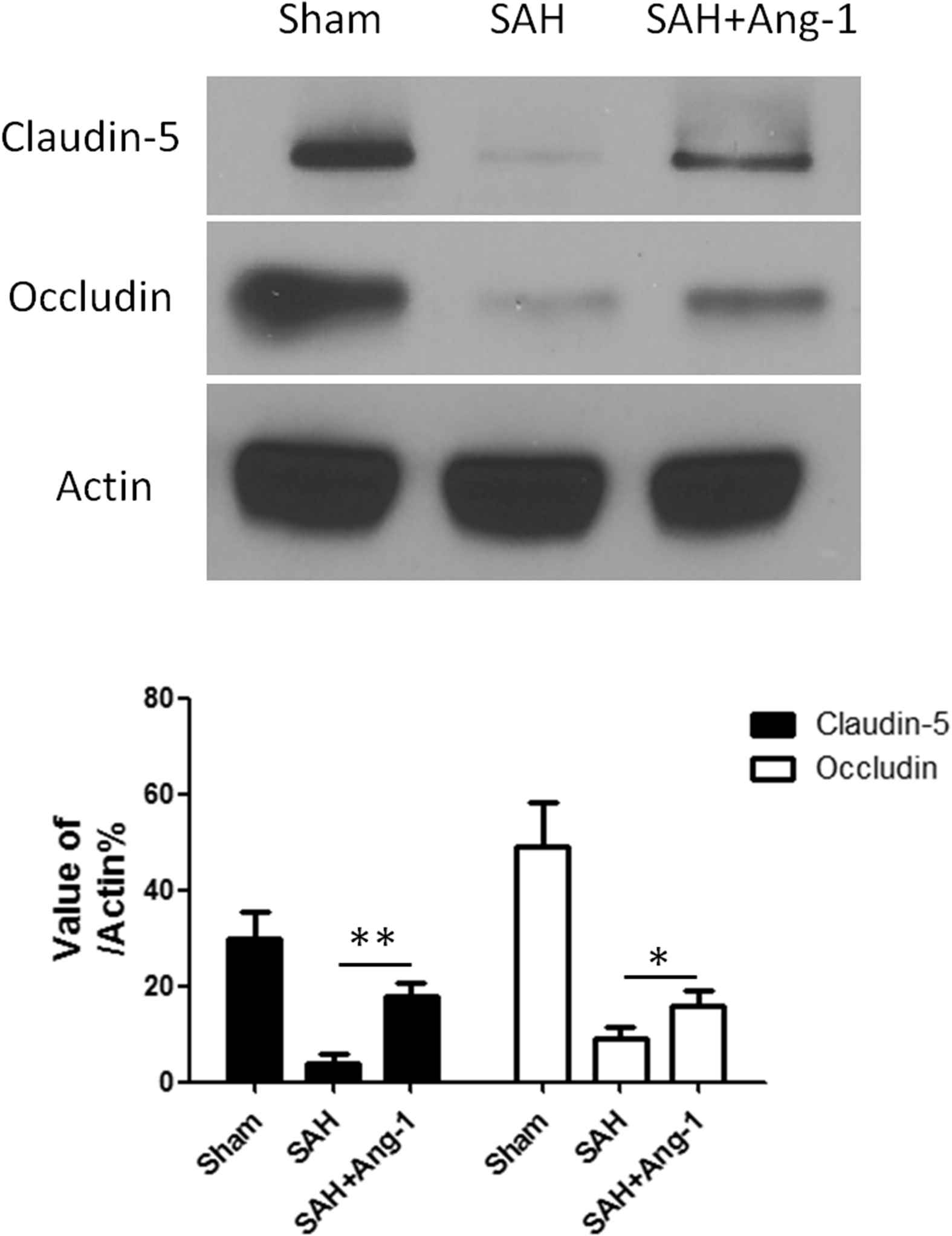
Western blot and quantitative analysis of Claudin-5 and Occludin in hippocampus. Results are described as mean ± standard deviation. Ang-1, angiopoietin-1; SAH, subarachnoid hemorrhage. *P < 0.05, **P < 0.01. versus the SAH group.
Our western blot analysis of pTie-2/Tie-2 have demonstrated that exogenous Ang-1 treatment increased the Tie-2 receptor phosphorylation level after SAH as described above. Furthermore, exogenous Ang-1 also improved the expression of Claudin-5 and Occludin in microvascular endothelial cells after SAH (Fig. 4). These manifestations were more pronounced in the hippocampus vascular area. The western blot analysis of Claudin-5 and Occludin in hippocampus confirmed the above results (Fig. 4).
Our published preliminary results have confirmed that Ang-1 expression decreases rapidly within 24 h after SAH.14 So this result indicated that the decline of Ang-1 expression may reduces the tight junction protein expression by decreasing the phosphorylation level of Tie-2 receptor.
Exogenous Ang-1 reduced BBB permeability after SAH
We did Evans blue extravasation at 24 h after SAH. Within 24 h after SAH, significant leakage of blood–brain barrier occurred. Evans blue viability assay showed that less Evans blue dye leaked in Ang-1 treatment group than SAH group (Fig. 5).
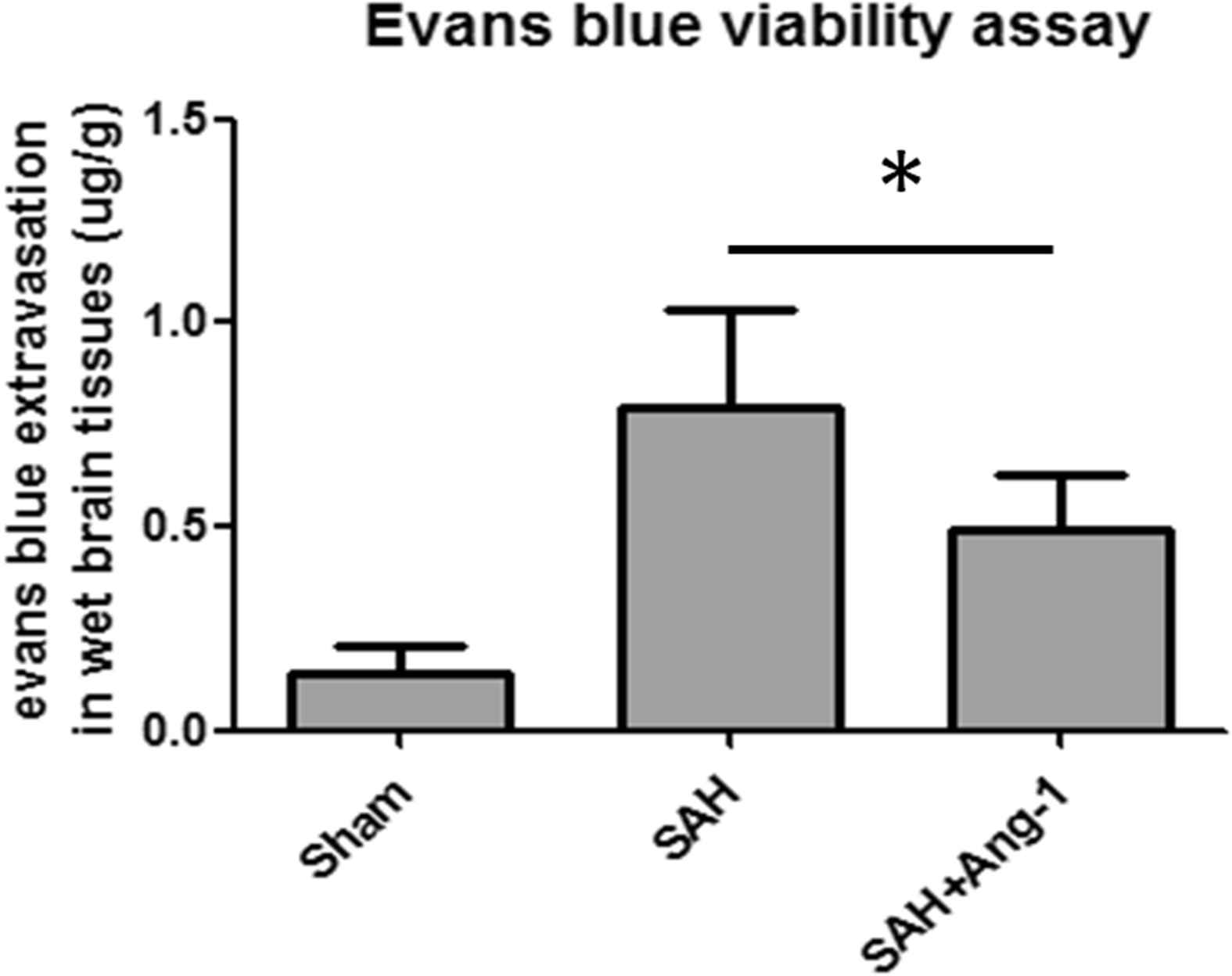
Evans blue extravasation quantitative analysis. Evans blue extravasation was evaluated at 24h after SAH. Results are described as mean ± standard deviation. Ang-1, angiopoietin-1; SAH, subarachnoid hemorrhage. *P < 0.05, **P < 0.01. versus the SAH group.
Discussion
Nowadays, more and more basic and clinical studies regard early brain injury (EBI) as the main research content and direction of SAH, and blood–brain barrier integrity is one of the research hotspots.15 Numerous studies have provided evidence that BBB disrupts in EBI.16 However, Cerebral vasospasm (CVS) was once considered of the main reason to lead to poor prognosis of survival after SAH, and has become a hotspot of clinical and basic researches. Unfortunately, the results of years of research shows, only Nimodipine is recognized as a clear improvement in the prognosis of clinical patients, but does not depend on its mitigation of CVS.17 In addition, recent clinical studies have found that the use of endothelin receptor antagonist Clazosentan, although able to effectively reduce the patient CVS, but does not improve the prognosis of patients.18 Since endothelin-1 receptor is located in smooth muscle cells, the experimental results from Nimodipine and Clazosentan suggest that not the arterial, which has the intact smooth muscle layer, but the cerebral microvascular, which is lack of smooth muscle layer, is one of the main factors of poor prognosis.
Our past research has focused on changes in microvascular endothelial cells in the early stages after SAH. Loosening of tight junctions of endothelial cells has been proved as an important initiating factor of early brain injury.7,8 Under physiological conditions, hyperphosphorylation of tie-2 receptors by high concentrations of Ang-1 is where brain microvascular endothelial cells differ from others. Our previous study has demonstrated that Ang-1 rapidly decreases after subarachnoid hemorrhage and appears at a lowest level at 24h. So we hoped to further understand the changes of tie-2 receptor phosphorylation and its effect on cerebral microvascular endothelial cells after SAH. So we designed this research and selected 24h after SAH as the time point.
In this study, we still chose internal carotid artery puncture model to simulate the process of subarachnoid hemorrhage. Its similarity with the rupture process of intracranial aneurysms is the unique advantage over subarachnoid blood injection model.19 Skilled and gentle microsurgical procedures avoided the high mortality rate, which was once the biggest drawback of endovascular perforation model.
Our results suggest that the Angiopoietin/Tie-2 signal path is involved in early brain injury after SAH. First, we successfully located the main region of the tie-2 receptor in rat brain tissue. The tie-2 receptor was mainly gathered in the area rich in blood vessels, particularly the hippocampus hypervascular region. Then we observed a significant decrease in the level of tie-2 receptor phosphorylation at 24h after SAH. Concurrently, the expression of tight junction protein (claudin-5 and occludin) in endothelial cells was down-regulated. Both of these changes could be corrected by the addition of exogenous Ang-1. Based on these results, we have every reason to believe that the phosphorylation of tie-2 receptors in brain microvascular endothelial cells decreased rapidly after SAH, and resulted in a lack of tight junction protein expression and blood–brain barrier leakage.
At the moment of the failure of CVS targeting treatment, a shift in treatment strategy is required urgently. Due to the existence of the blood–brain barrier, the current administrations of the central nervous system have apparent limitations. Intraventricular injection and dural infiltration administration have the disadvantages of being invasive and complicated. The therapy targeting the microvascular endothelial cells do not need penetrating the blood–brain barrier, and should be considered as one of the future feasible treatment strategies of SAH. Our research provides a possible therapeutic approach: restore the integrity of blood–brain barrier by increasing tie-2 receptor phosphorylation. We look forward to seeing more relevant basic and clinical research in the near future.
Conflict of interest
No conflict of interest exits in the submission of this manuscript, and manuscript is approved by all authors for publication.
References
Cite this article
TY - JOUR AU - Hua Gu AU - Yi-qi Wang AU - Chao-Hui Zhao AU - Xing-ming Zhong AU - Jian-guo Yang PY - 2018 DA - 2018/08/23 TI - The decrease of Tie-2 receptor phosphorylation in microvascular endothelial cells is involved in early brain injury after subarachnoid hemorrhage JO - Artery Research SP - 45 EP - 51 VL - 23 IS - C SN - 1876-4401 UR - https://doi.org/10.1016/j.artres.2018.07.004 DO - 10.1016/j.artres.2018.07.004 ID - Gu2018 ER -